条鰭類
条鰭類(じょうきるい、Actinopterygii)は、魚類(=四肢動物以外の脊椎動物)の下位分類群の一つ[3]。分類階級としては条鰭亜綱とする場合、条鰭綱とする場合、条鰭上綱とする場合がある。種数では現存する脊椎動物の半数以上を占める[4]。肉鰭類の肉厚な葉状の鰭とは対照的に、放射状に伸びる細い鰭条から支えられる鰭を持つ。鰭は扇子のように畳んだり開いたりすることができ、軟骨魚類や肉鰭類と比べて鰭の形状や面積を自由に変えられる。これにより推力重量比が優れたものとなる。鰭条は橈骨と接続し、鰭と内部器官を結合している。
| 条鰭亜綱 | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 | ||||||||||||||||||
| 分類 | ||||||||||||||||||
| ||||||||||||||||||
| 学名 | ||||||||||||||||||
| Actinopterygii Klein, 1885[2] |
条鰭類の大半は真骨類である。条鰭類は脊椎動物の大部分を占め、現存する3万種を超える魚類の約95%を占める[5]。最も数の多い水生動物であり、深海や地下から、最も標高の高い山の渓流まで、淡水と海洋のあらゆる環境に遍在する。体長8 mmのパエドキプリス・プロゲネティカから、体重2,300 kgの巨大なウシマンボウ、全長11 mのリュウグウノツカイまで様々である。これまで知られている最大の条鰭類は、ジュラ紀のリードシクティスで、16.5 mまで成長したと推定されている。
形態
編集
条鰭類には多くの特徴的な形態を持った種が分類される。典型的な条鰭類の主な特徴を隣の図に示す。
鰾はより派生した構造で、浮力維持に使用される[6]。肉鰭類の肺と同様に前腸の腹側から発達するという祖先の状態を保持しているポリプテルス類を除き、条鰭類の鰾は前腸の背側から発達する[6][7]。初期の形態では、鰾はまだ呼吸に使用されており、この特徴は全骨類(アミア類とガー類)に残されている[8]。ピラルクなどの一部の魚では、鰾は再び空気呼吸のために変化しているが[9]、他の系統では空気呼吸の機能は完全に失われている[10]。
鰭類の骨格は一部の原始的な分類群を除き、ほぼ完全に硬骨によって構成されている[11]。鱗の形態は硬鱗、円鱗あるいは櫛鱗など多様で、鱗をもたないグループも多い。鱗のある真骨類はすべて葉状鱗を持つ。これらの鱗の外側は骨隆起とともに扇状に広がり、内側は繊維状の結合組織で交差している。葉状鱗は他の種類の鱗よりも薄く透明で、硬化したエナメル質や象牙質のような層がない。真骨類以外の条鰭類に見られるガノイン鱗とは異なり、成長するにつれて同心円状に鱗が大きくなる[12]。
真骨類と軟質類は、古倍数性によるゲノム重複を経験したという点で、多鰭類と全骨類とは異なる。ゲノム重複は、平均して遺伝子重複の約17%を保持している真骨類で約3億2000万年前に発生し、軟質類では約1億8000万年前(1億2400万 - 2億2500万年前)に発生したと推定されている。その後、サケ科(8000万 - 1億年前)などの一部の系統で再び発生し、コイ科(1400万年前と最近)内でも独立して数回発生している[13][14][15][16][17]。
鰭は担鰭骨に支えられる鰭条と、鰭条同士をつなぐ鰭膜によって構成される。ポリプテルス目を除き、胸鰭の射出骨は肩甲骨・烏口骨複合体と接続する。ほとんどの仲間は間鰓蓋骨と鰓条骨をもつ[18]。咽頭板を欠き、鼻孔は頭部の比較的上方に位置し内鼻孔をもたない[19]。
形態の多様性
編集条鰭類には様々な形態があり、ここではその一部を挙げる。
-
 マグロは速く泳ぐために流線型の体と二叉する尾鰭をもつ
マグロは速く泳ぐために流線型の体と二叉する尾鰭をもつ -
 メカジキはマグロよりも流線型の体をしている
メカジキはマグロよりも流線型の体をしている -
 サケは尾の推進力で川を遡上する
サケは尾の推進力で川を遡上する -
 タラには3基の背鰭と2基の臀鰭があり、機動力が高い
タラには3基の背鰭と2基の臀鰭があり、機動力が高い -
 カレイ目は背鰭と臀鰭が発達している
カレイ目は背鰭と臀鰭が発達している -
 ヨツメウオは目が二つに分かれ、水上と水中の両方を見られる
ヨツメウオは目が二つに分かれ、水上と水中の両方を見られる -
 ハダカイワシは腹に発光器がある
ハダカイワシは腹に発光器がある -
 ヨコエソ科も発光器があり、口には剛毛のような歯がある
ヨコエソ科も発光器があり、口には剛毛のような歯がある -
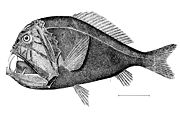 オニキンメは待ち伏せ型の捕食者である
オニキンメは待ち伏せ型の捕食者である -
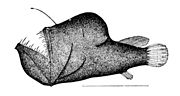 アンコウ目の背鰭棘は釣り竿のような誘引突起に変化している
アンコウ目の背鰭棘は釣り竿のような誘引突起に変化している -
 キンメダイは光を逃さないため眼が大きい
キンメダイは光を逃さないため眼が大きい -
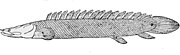 ポリプテルスは原始的な条鰭類で、肺もある
ポリプテルスは原始的な条鰭類で、肺もある -
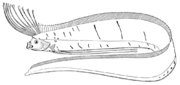 リュウグウノツカイは最長の条鰭類である
リュウグウノツカイは最長の条鰭類である -
 ヨーロッパアナゴは細長い体形をしている
ヨーロッパアナゴは細長い体形をしている -
 ミノカサゴ属は鰭条に毒を持つ
ミノカサゴ属は鰭条に毒を持つ -
 フウリュウウオは胸鰭で海底を歩き回る
フウリュウウオは胸鰭で海底を歩き回る -
 フウセンウナギは胃がとても大きい
フウセンウナギは胃がとても大きい -
 モルミルス科の一部は下顎が発達する
モルミルス科の一部は下顎が発達する -
 チョウザメ科は一部が軟骨である
チョウザメ科は一部が軟骨である -
 ダツ科は細長く尖った吻を持つ
ダツ科は細長く尖った吻を持つ -
 タツノオトシゴは特異な形態を持つ
タツノオトシゴは特異な形態を持つ -
 カガミダイはその名の通り銀色の体を持つ
カガミダイはその名の通り銀色の体を持つ -
 シイラは頭部が発達する
シイラは頭部が発達する -
 トビウオ科は胸鰭で海上を滑空する
トビウオ科は胸鰭で海上を滑空する -
 マンボウ科は尾鰭が無い
マンボウ科は尾鰭が無い -
 歴史上最大の条鰭類、リードシクティス
歴史上最大の条鰭類、リードシクティス -
 ハコフグ科は固い骨板で覆われる
ハコフグ科は固い骨板で覆われる -
 ガー目とアミア目で全骨類を構成する
ガー目とアミア目で全骨類を構成する

.png)









.jpg)





.jpg)









繁殖
編集
多くの種が雌雄異体であり、雌は卵を産んで、卵は体外で受精する種が多い。通常、産まれた卵は雄によって受精される。その後、自由に泳ぐ仔魚を経て発育が進む[20]。雌雄同体の種もいる。ほとんどの場合雌性先熟であり、雌として生まれ、ある段階で雄に変化する。雄から雌に変化する雄性先熟は、雌性先熟よりも少ない[21]。
ほとんどの科は、体内受精ではなく体外受精を行う[22]。卵生の真骨類のうち79%は親による育児を行わない[23]。胎生、卵胎生、または親による卵の育児は、422の真骨類の科のうち、21%で見られる。育児を行わないのが祖先の状態である可能性が高い[23]。条鰭類における胎生の最古の例は、中期三畳紀のサウリクティスで発見されている[24]。胎生は比較的まれで、現生の真骨類の約6%に見られる。雄による育児は雌による育児よりもはるかに一般的である[23][25]。雄の縄張り意識が、進化の過程で育児に発達した可能性がある[26][27]。
自家受精する例はいくつかある。マングローブ・キリフィッシュは両性具有で、体内受精をする。この繁殖方法は、生息するマングローブ林で長期間水から出るという習性に関係している可能性がある。雄は19℃以下の温度で生まれることがあり、雌が産卵させる卵を受精させることができる。これにより、近親交配する種でも遺伝的多様性が維持される[28]。
分類と化石記録
編集
姉妹群は四肢動物、ハイギョ、シーラカンスからなる肉鰭亜綱で、肉鰭亜綱と条鰭亜綱を合わせて硬骨魚類と総称される。条鰭類は腕鰭類とActinopteriに分けられる。Actinopteriは軟質類と新鰭類を含む。新鰭類はさらに全骨類と真骨類に分けられる。中生代(三畳紀、ジュラ紀、白亜紀)と新生代には特に真骨類が大きく多様化した。その結果、現生魚類の96%が真骨類(全魚種の40%が真骨類内のAcanthomorphaに属する)であり、その他の条鰭類は多様化していない[29]。最古の条鰭類は古生代のシルル紀後期に出現したとみられ、Andreolepis 属など5属が知られている。続くデボン紀から中生代三畳紀にかけて栄えた軟質亜綱の仲間は、ジュラ紀終盤までにチョウザメ目を残しほとんどが絶滅している。白亜紀以降は、高い運動能力と効率的な摂餌機構を発達させた条鰭類のサブグループである新鰭類が支配的な地位を獲得し、水圏のあらゆる環境に適応放散を果たした[30]。新鰭類の魚類は、現代では約2万6800種を擁する脊椎動物の中で最大のグループとなっている[30]。
下の系統図は、現生条鰭類の主な系統群と、他の現生魚類および四肢動物との進化的関係を示している[31][32][33][34]。四肢動物には主に陸生種が含まれるが、二次的に水生になった種(クジラやイルカなど)も含まれる。四肢動物はデボン紀に硬骨魚類のグループから進化した[35]。異なる条鰭綱の系統群のおおよその分岐日(百万年、mya)は、Near et al., 2012によるものである[33]。
| 脊椎動物 |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
ポリプテルス目はその他の条鰭類の姉妹群であり、チョウザメ目は新鰭類の姉妹群であり、全骨類は真骨類の姉妹群である。カライワシ上目は最も原始的な真骨類であると考えられている[33]。
最も古い化石条鰭類はアンドレオレピスで、4億2千万年前のシルル紀後期の地層から、ロシア、スウェーデン、エストニアで化石が発見されている[36]。条鰭類のクラウングループは、おそらくデボン紀と石炭紀の境界付近に起源を持つ[37]。現代の真骨類の最も古い化石近縁種は三畳紀のものであるが[38][39]、真骨類は古生代にはすでに起源を持っていたと疑われている[33]。
| 軟質類 | 軟質類は骨格の一部が軟骨になっている。以前の軟質類は側系統群であり、共通祖先の子孫がすべて含まれているわけではない。かつては52種がチョウザメ目とポリプテルス目に分かれていた。現在ポリプテルス目は独自の腕鰭類とされる。軟質類は硬骨魚類から進化したが、その過程で軟骨部分の骨化が失われた。高齢個体では骨化が進むため、軟質類では骨化が失われたのではなく、遅れていることが示唆されている[40]。軟質類は軟骨があり、顎の構造もサメに似ており、鱗が無いことから、サメとされることもあった。また噴水孔があり、尾も異尾である。しかし化石記録からは、外見よりも真骨類に近いことが示されている[40]。 | |
|---|---|---|
| 新鰭類 | 新鰭類は後期ペルム紀に出現した。進化の過程で、それ以前の条鰭類からの変化はわずかであった。新鰭類は祖先よりも素早く移動できるため、非常に成功したグループである。進化の過程で鱗と骨格が軽くなり、顎はより強力で効率的になった。ロレンチーニ器官はヌタウナギを除く他のすべての魚類グループに存在するが、新鰭類はこれを失っている。後にデンキウナギ目とナマズ目で再進化した[41]。 |












.jpg)




下位分類
編集以下に絶滅分類群を含む分類群を、系統順位に沿って目の単位まで示す。各グループの詳細、内部に含まれる絶滅群については、それぞれの項目を参照。分類はBetancur-R, Ricardo; et al. (2013).、Betancur-Rodriguez; et al. (2017).を参考[34][42]。また、Nelson[4]、ITIS[43]、FishBase[44]も参考。絶滅分類群はVan der Laan (2016)[45]、Xu (2021)[46]を参考。
- †?Asarotiformes Schaeffer, 1968
- †?Discordichthyiformes Minikh, 1998
- †?Paphosisciformes Grogan & Lund, 2015
- †?Scanilepiformes Selezneya, 1985 スカニレピス目
- †Cheirolepidiformes Kazantseva-Selezneva, 1977 ケイロレピス目
- †Paramblypteriformes Heyler, 1969 パラムブリプテルス目
- †Rhadinichthyiformes Lowney, 1980
- †Palaeonisciformes Hay, 1902 パレオニスクム目
- †Tarrasiiformes sensu Lund & Poplin, 2002 タッラシウス目
- †Ptycholepiformes Andrews et al. 1967
- †Haplolepidiformes Westoll, 1944
- †Aeduelliformes Heyler, 1969 アエデュエラ目
- †Platysomiformes Aldinger, 1937 プラティソムス目
- †Dorypteriformes Cope, 1871 ドリプテルス目
- †Eurynotiformes Sallan & Coates, 2013 ユーリノトゥス目
- 腕鰭類(多鰭類、分岐鰭類) Cladistia Pander, 1860
- †Guildayichthyiformes Lund, 2000 グイルダイクティス目
- Polypteriformes Bleeker, 1859 ポリプテルス目
- Actinopteri Cope, 1972 s.s.
- †Elonichthyiformes Kazantseva-Selezneva, 1977 エロニクティス目
- †Phanerorhynchiformes Long, 2011 ファネロリンクス目
- †Bobasatraniiformes Berg, 1940 ボバサトラニア目
- †Saurichthyiformes Aldinger, 1937 サウリクティス目
- 軟質類 Chondrostei Müller, 1844
- †Birgeriiformes Heyler, 1969 ビルゲリア目
- †Chondrosteiformes Aldinger, 1937 コンドロステウス目
- Acipenseriformes Berg, 1940 チョウザメ目
- 新鰭類 Neopterygii Regan, 1923 sensu Xu & Wu, 2012
- †Pholidopleuriformes Berg, 1937 フォリドプレウルス目
- †Redfieldiiformes Berg, 1940
- †Platysiagiformes Brough, 1939
- †Polzbergiiformes Griffith, 1977
- †Perleidiformes Berg, 1937 ペルレイドゥス目
- †Louwoichthyiformes Xu, 2021
- †Peltopleuriformes Lehman, 1966 ペルトプレウルス目
- †Luganoiiformes Lehman, 1958 ルガノイア目
- †Pycnodontiformes Berg, 1937 ピクノドン目
- 全骨類 Holostei Müller, 1844
- Halecomorphi Cope, 1872 sensu Grande & Bemis, 1998
- †Parasemionotiformes Lehman, 1966 パラセミオノトゥス目
- †Ionoscopiformes Grande & Bemis, 1998 イオノスコプス目
- Amiiformes Huxley, 1861 sensu Grande & Bemis, 1998 アミア目
- Ginglymodi Cope, 1871
- †Dapediiformes Thies & Waschkewitz, 2015 ダペディウム目
- †Semionotiformes Arambourg & Bertin, 1958 セミオノトゥス目
- Lepisosteiformes Hay, 1929 ガー目
- Halecomorphi Cope, 1872 sensu Grande & Bemis, 1998
- Teleosteomorpha Arratia, 2000 sensu Arratia, 2013
- †Prohaleciteiformes Arratia, 2017
- Aspidorhynchei Nelson, Grand & Wilson, 2016 アスピドリンクス亜区
- †Aspidorhynchiformes Bleeker, 1859 アスピドリンクス目
- †Pachycormiformes Berg, 1937 パキコルムス目
- 真骨類 Teleostei Müller, 1844 sensu Arratia, 2013
- †?Araripichthyiformes Nelson et al., 2016 アラリピクティス目
- †?Ligulelliformes Taverne, 2011
- †?Tselfatiiformes Nelson, 1994 ツェルファティア目
- †Pholidophoriformes Berg, 1940 フォリドフォルス目
- †Dorsetichthyiformes Nelson, Grand & Wilson, 2016
- †Leptolepidiformes レプトレピス目
- †Crossognathiformes Taverne, 1989 クロッソグナトゥス目
- †Ichthyodectiformes Bardeck & Sprinkle, 1969 イクチオデクテス目
- Teleocephala de Pinna, 1996 s.s.
- Elopocephalai カライワシ巨区 Patterson, 1977 sensu Arratia, 1999 (Elopomorpha Greenwood et al., 1966 カライワシ上目)
- Elopiformes Gosline, 1960 カライワシ目
- Albuliformes Greenwood et al., 1966 sensu Forey et al., 1996 ソトイワシ目
- Notacanthiformes Goodrich, 1909 ソコギス目
- Anguilliformes Jarocki, 1822 sensu Goodrich, 1909 ウナギ目
- Osteoglossocephalai アロワナ巨区 sensu Arratia 1999
- Osteoglossocephala アロワナ上区 sensu Arratia, 1999 (Osteoglossomorpha Greenwood et al., 1966 アロワナ上目)
- †Lycopteriformes Chang & Chou, 1977 リコプテラ目
- Hiodontiformes McAllister, 1968 sensu Taverne, 1979 ヒオドン目
- Osteoglossiformes Regan, 1909 sensu Zhang, 2004 アロワナ目
- Clupeocephala ニシン上目 Patterson & Rosen, 1977 sensu Arratia, 2010
- Otomorpha 骨鰾区 Wiley & Johnson, 2010 (Otocephala; Ostarioclupeomorpha 骨鰾上目)
- Clupei ニシン亜区 Wiley & Johnson, 2010 (Clupeomorpha Greenwood et al., 1966 ニシン上目)
- †Ellimmichthyiformes Grande, 1982
- Clupeiformes Bleeker, 1859 ニシン目
- Alepocephali
- Alepocephaliformes Marshall, 1962 セキトリイワシ目
- Ostariophysi 骨鰾亜区 Sagemehl, 1885
- Anotophysa (Rosen & Greenwood, 1970) Sagemehl, 1885
- †Sorbininardiformes Taverne, 1999
- Gonorynchiformes Regan, 1909 ネズミギス目
- Otophysa Garstang, 1931
- Cypriniformes Bleeker, 1859 sensu Goodrich, 1909 コイ目
- Characiformes Goodrich, 1909 カラシン目
- Gymnotiformes Berg, 1940 デンキウナギ目
- Siluriformes Cuvier, 1817 sensu Hay 1929 ナマズ目
- Anotophysa (Rosen & Greenwood, 1970) Sagemehl, 1885
- Clupei ニシン亜区 Wiley & Johnson, 2010 (Clupeomorpha Greenwood et al., 1966 ニシン上目)
- Euteleosteomorpha (Greenwood et al., 1966) (Euteleostei Greenwood, 1967 sensu Johnson & Patterson, 1996 正真骨類)
- Lepidogalaxii
- Lepidogalaxiiformes Betancur-Rodriguez et al., 2013 レピドガラクシアス目
- Protacanthopterygii 原棘鰭上目 Greenwood et al., 1966 sensu Johnson & Patterson, 1996
- Argentiniformes ニギス目
- Galaxiiformes ガラクシアス目
- Salmoniformes Bleeker, 1859 sensu Nelson, 1994 サケ目
- Esociformes Bleeker, 1859 カワカマス目
- Stomiati
- Osmeriformes キュウリウオ目
- Stomiatiformes Regan, 1909 ワニトカゲギス目
- Neoteleostei 新真骨類 Nelson, 1969
- Ateleopodia
- Eurypterygia Rosen, 1973
- Aulopa [Cyclosquamata Rosen, 1973 円鱗上目]
- Aulopiformes Rosen, 1973 ヒメ目
- Ctenosquamata Rosen, 1973
- Myctophata [Scopelomorpha]
- Myctophiformes Regan, 1911 ハダカイワシ目
- Acanthomorpha Betancur-Rodriguez et al., 2013
- Lampridacea Betancur-Rodriguez et al., 2013 [Lampridomorpha; Lampripterygii アカマンボウ上目]
- Lampriformes Regan, 1909 アカマンボウ目
- Paracanthomorphacea sensu Grande et al., 2013 (Paracanthopterygii 側棘鰭上目 Greenwood, 1937)
- Percopsiformes Berg, 1937 サケスズキ目
- †Sphenocephaliformes Rosen & Patterson, 1969
- Zeiformes Regan, 1909 マトウダイ目
- Stylephoriformes Miya et al., 2007 ステューレポルス目
- Gadiformes Goodrich, 1909 タラ目
- Polymixiacea Betancur-Rodriguez et al., 2013 (Polymyxiomorpha; Polymixiipterygii ギンメダイ上目)
- †Pattersonichthyiformes Gaudant, 1976
- †Ctenothrissiformes Berg, 1937 クテノトリッサ目
- Polymixiiformes Lowe, 1838 ギンメダイ目
- Euacanthomorphacea Betancur-Rodriguez et al., 2013 (Euacanthomorpha sensu Johnson & Patterson, 1993; Acanthopterygii 棘鰭上目 Gouan, 1770 sensu])
- Berycimorphaceae Betancur-Rodriguez et al., 2013
- Holocentrimorphaceae Betancur-Rodriguez et al., 2013
- Percomorphaceae Betancur-Rodriguez et al., 2013 (Percomorpha スズキ系 sensu Miya et al., 2003; Acanthopteri)
- Ophidiimopharia Betancur-Rodriguez et al., 2013
- Batrachoidimopharia Betancur-Rodriguez et al., 2013
- Gobiomopharia Betancur-Rodriguez et al., 2013
- Scombrimopharia Betancur-Rodriguez et al., 2013
- Carangimopharia Betancur-Rodriguez et al., 2013
- Anabantaria Betancur-Rodriguez et al., 2014
- Carangaria Betancur-Rodriguez et al., 2014
- Carangaria incertae sedis
- Istiophoriformes Betancur-Rodriguez, 2013 カジキ目
- Carangiformes アジ目
- Pleuronectiformes Bleeker, 1859 カレイ目
- Ovalentaria オヴァレンタリア類 Smith & Near, 2012 (Stiassnyiformes sensu Li et al., 2009)
- Ovalentaria incertae sedis
- Cichliformes Betancur-Rodriguez et al., 2013 カワスズメ目(ポリディクテュス科、ポリケントルス科を含む)
- Atheriniformes Rosen, 1964 トウゴロウイワシ目
- Cyprinodontiformes Berg, 1940 カダヤシ目
- Beloniformes Berg, 1940 ダツ目
- Mugiliformes Berg, 1940 ボラ目
- Blenniiformes Springer, 1993 イソギンポ目
- Gobiesociformes Gill, 1872 ウバウオ目
- Eupercaria ユーペルカ類 Betancur-Rodriguez et al., 2014 (Percomorpharia Betancur-Rodriguez et al., 2013)
- Eupercaria incertae sedis
- Gerreiformes クロサギ目
- Labriformes ベラ目(ブダイ科を含む)
- Caproiformes ヒシダイ目
- Lophiiformes Garman, 1899 アンコウ目
- Tetraodontiformes Regan, 1929 フグ目
- Centrarchiformes Bleeker, 1859 サンフィッシュ目
- Gasterosteiformes トゲウオ目
- Scorpaeniformes カサゴ目
- Perciformes Bleeker, 1859 スズキ目
- Lampridacea Betancur-Rodriguez et al., 2013 [Lampridomorpha; Lampripterygii アカマンボウ上目]
- Myctophata [Scopelomorpha]
- Aulopa [Cyclosquamata Rosen, 1973 円鱗上目]
- Lepidogalaxii
- Otomorpha 骨鰾区 Wiley & Johnson, 2010 (Otocephala; Ostarioclupeomorpha 骨鰾上目)
- Osteoglossocephala アロワナ上区 sensu Arratia, 1999 (Osteoglossomorpha Greenwood et al., 1966 アロワナ上目)
- Elopocephalai カライワシ巨区 Patterson, 1977 sensu Arratia, 1999 (Elopomorpha Greenwood et al., 1966 カライワシ上目)
出典・脚注
編集- ^ Zhao, W.; Zhang, X.; Jia, G.; Shen, Y.; Zhu, M. (2021). “The Silurian-Devonian boundary in East Yunnan (South China) and the minimum constraint for the lungfish-tetrapod split”. Science China Earth Sciences 64 (10): 1784–1797. Bibcode: 2021ScChD..64.1784Z. doi:10.1007/s11430-020-9794-8.
- ^ 資料によって一定しない。[1] によると他に Cope, 1871, Cope, 1887, Cope, 1891, Woodward, 1891 など。[2] は(理由を挙げずに)「Cope, 1887 ではなく Klein, 1885」としている。
- ^ Kardong, Kenneth (2015). Vertebrates: Comparative Anatomy, Function, Evolution. New York: McGraw-Hill Education. pp. 99–100. ISBN 978-0-07-802302-6
- ^ a b Nelson, Joseph S. (2016). Fishes of the World. John Wiley & Sons. ISBN 978-1-118-34233-6
- ^ (Davis, Brian 2010).
- ^ a b Funk, Emily; Breen, Catriona; Sanketi, Bhargav; Kurpios, Natasza; McCune, Amy (2020). “Changing in Nkx2.1, Sox2, Bmp4, and Bmp16 expression underlying the lung-to-gas bladder evolutionary transition in ray-finned fishes”. Evolution & Development 22 (5): 384–402. doi:10.1111/ede.12354. PMC 8013215. PMID 33463017.
- ^ Funk, Emily C.; Breen, Catriona; Sanketi, Bhargav D.; Kurpios, Natasza; McCune, Amy (25 September 2020). “Changes in Nkx2.1, Sox2, Bmp4, and Bmp16 expression underlying the lung-to-gas bladder evolutionary transition in ray-finned fishes”. Evolution & Development 22 (5): 384–402. doi:10.1111/ede.12354. PMC 8013215. PMID 33463017.
- ^ Zhang, Ruihua; Liu, Qun; Pan, Shanshan; Zhang, Yingying; Qin, Yating; Du, Xiao; Yuan, Zengbao; Lu, Yongrui et al. (13 September 2023). “A single-cell atlas of West African lungfish respiratory system reveals evolutionary adaptations to terrestrialization”. Nature Communications 14 (1): 5630. Bibcode: 2023NatCo..14.5630Z. doi:10.1038/s41467-023-41309-3. PMC 10497629. PMID 37699889.
- ^ Scadeng, Miriam; McKenzie, Christina; He, Weston; Bartsch, Hauke; Dubowitz, David J.; Stec, Dominik; St. Leger, Judy (25 November 2020). “Morphology of the Amazonian Teleost Genus Arapaima Using Advanced 3D Imaging”. Frontiers in Physiology 11: 260. doi:10.3389/fphys.2020.00260. PMC 7197331. PMID 32395105.
- ^ Martin, Rene P; Dias, Abigail S; Summers, Adam P; Gerringer, Mackenzie E (16 October 2022). “Bone Density Variation in Rattails (Macrouridae, Gadiformes): Buoyancy, Depth, Body Size, and Feeding”. Integrative Organismal Biology 4 (1): obac044. doi:10.1093/iob/obac044. PMC 9652093. PMID 36381998.
- ^ 『魚学入門』 p.23
- ^ “Actinopterygii Klein, 1885” (英語). www.gbif.org. 2024年6月17日閲覧。
- ^ Davesne, Donald; Friedman, Matt; Schmitt, Armin D.; Fernandez, Vincent; Carnevale, Giorgio; Ahlberg, Per E.; Sanchez, Sophie; Benson, Roger B. J. (27 July 2021). “Fossilized cell structures identify an ancient origin for the teleost whole-genome duplication”. Proceedings of the National Academy of Sciences 118 (30). Bibcode: 2021PNAS..11801780D. doi:10.1073/pnas.2101780118. PMC 8325350. PMID 34301898.
- ^ Parey, Elise; Louis, Alexandra; Montfort, Jerome; Guiguen, Yann; Crollius, Hugues Roest; Berthelot, Camille (12 August 2022). “An atlas of fish genome evolution reveals delayed rediploidization following the teleost whole-genome duplication”. Genome Research 32 (9): 1685–1697. doi:10.1101/gr.276953.122. PMC 9528989. PMID 35961774.
- ^ Du, Kang; Stöck, Matthias; Kneitz, Susanne; Klopp, Christophe; Woltering, Joost M.; Adolfi, Mateus Contar; Feron, Romain; Prokopov, Dmitry et al. (2020). “The sterlet sturgeon genome sequence and the mechanisms of segmental rediploidization” (英語). Nature Ecology & Evolution 4 (6): 841–852. Bibcode: 2020NatEE...4..841D. doi:10.1038/s41559-020-1166-x. ISSN 2397-334X. PMID 32231327.
- ^ Kuraku, Shigehiro; Sato, Mana; Yoshida, Kohta; Uno, Yoshinobu (2024). “Genomic reconsideration of fish non-monophyly: why cannot we simply call them all 'fish'?” (英語). Ichthyological Research 71 (1): 1–12. Bibcode: 2024IchtR..71....1K. doi:10.1007/s10228-023-00939-9. ISSN 1616-3915.
- ^ Xu, Peng; Xu, Jian; Liu, Guangjian; Chen, Lin; Zhou, Zhixiong; Peng, Wenzhu; Jiang, Yanliang; Zhao, Zixia et al. (2019). “The allotetraploid origin and asymmetrical genome evolution of the common carp Cyprinus carpio” (英語). Nature Communications 10 (1): 4625. Bibcode: 2019NatCo..10.4625X. doi:10.1038/s41467-019-12644-1. ISSN 2041-1723. PMID 31604932.
- ^ 分岐鰭亜綱は鰓条骨を、軟質下綱は間鰓蓋骨をそれぞれ欠く。
- ^ 『Fishes of the World Fourth Edition』 pp.87–88
- ^ Dorit, R.L.; Walker, W.F.; Barnes, R.D. (1991). Zoology. Saunders College Publishing. p. 819. ISBN 978-0-03-030504-7
- ^ Avise, J.C.; Mank, J.E. (2009). “Evolutionary perspectives on hermaphroditism in fishes”. Sexual Development 3 (2–3): 152–163. doi:10.1159/000223079. PMID 19684459.
- ^ Pitcher, T (1993). The Behavior of Teleost Fishes. London: Chapman & Hall
- ^ a b c Reynolds, John; Nicholas B. Goodwin; Robert P. Freckleton (19 March 2002). “Evolutionary Transitions in Parental Care and Live Bearing in Vertebrates”. Philosophical Transactions of the Royal Society B: Biological Sciences 357 (1419): 269–281. doi:10.1098/rstb.2001.0930. PMC 1692951. PMID 11958696.
- ^ Maxwell (2018). “Re-evaluation of the ontogeny and reproductive biology of the Triassic fish Saurichthys (Actinopterygii, Saurichthyidae)”. Palaeontology 61: 559–574. doi:10.5061/dryad.vc8h5.
- ^ Clutton-Brock, T. H.『The Evolution of Parental Care』Princeton UP、Princeton, NJ、1991年。
- ^ Werren, John; Mart R. Gross; Richard Shine (1980). “Paternity and the evolution of male parentage”. Journal of Theoretical Biology 82 (4): 619–631. doi:10.1016/0022-5193(80)90182-4. PMID 7382520 15 September 2013閲覧。.
- ^ Baylis, Jeffrey (1981). “The Evolution of Parental Care in Fishes, with reference to Darwin's rule of male sexual selection”. Environmental Biology of Fishes 6 (2): 223–251. Bibcode: 1981EnvBF...6..223B. doi:10.1007/BF00002788.
- ^ Wootton, Robert J.; Smith, Carl (2014). Reproductive Biology of Teleost Fishes. Wiley. ISBN 978-1-118-89139-1
- ^ Sallan, Lauren C. (February 2014). “Major issues in the origins of ray-finned fish (Actinopterygii) biodiversity”. Biological Reviews 89 (4): 950–971. doi:10.1111/brv.12086. hdl:2027.42/109271. PMID 24612207.
- ^ a b 『日本の海水魚』 pp.14-18
- ^ キャンベル11版 p.826.
- ^ 日本動物学会2018 pp.92-93
- ^ a b c d Thomas J. Near (2012). “Resolution of ray-finned fish phylogeny and timing of diversification”. PNAS 109 (34): 13698–13703. Bibcode: 2012PNAS..10913698N. doi:10.1073/pnas.1206625109. PMC 3427055. PMID 22869754.
- ^ a b Betancur-R, Ricardo (2013). “The Tree of Life and a New Classification of Bony Fishes”. PLOS Currents Tree of Life 5 (Edition 1). doi:10.1371/currents.tol.53ba26640df0ccaee75bb165c8c26288. hdl:2027.42/150563. PMC 3644299. PMID 23653398.
- ^ Laurin, M.; Reisz, R.R. (1995). “A reevaluation of early amniote phylogeny”. Zoological Journal of the Linnean Society 113 (2): 165–223. doi:10.1111/j.1096-3642.1995.tb00932.x.
- ^ “Fossilworks: Andreolepis”. 12 February 2010時点のオリジナルよりアーカイブ。14 May 2008閲覧。
- ^ Henderson, Struan; Dunne, Emma M.; Fasey, Sophie A.; Giles, Sam (3 October 2022). “The early diversification of ray-finned fishes (Actinopterygii): hypotheses, challenges and future prospects”. Biological Reviews 98 (1): 284–315. doi:10.1111/brv.12907. PMC 10091770. PMID 36192821.
- ^ Arratia, G. (2015). “Complexities of early teleostei and the evolution of particular morphological structures through time.”. Copeia 103 (4): 999–1025. doi:10.1643/CG-14-184.
- ^ Romano, Carlo; Koot, Martha B.; Kogan, Ilja; Brayard, Arnaud; Minikh, Alla V.; Brinkmann, Winand; Bucher, Hugo; Kriwet, Jürgen (February 2016). “Permian-Triassic Osteichthyes (bony fishes): diversity dynamics and body size evolution”. Biological Reviews 91 (1): 106–147. doi:10.1111/brv.12161. PMID 25431138.
- ^ a b “Chondrosteans: Sturgeon Relatives”. paleos.com. 25 December 2010時点のオリジナルよりアーカイブ。2024年6月17日閲覧。
- ^ Theodore Holmes Bullock、Carl D. Hopkins、Arthur N. Popper『Electroreception』Springer Science+Business Media, Incorporated、2005年、229頁。ISBN 978-0-387-28275-6。
- ^ Betancur-Rodriguez (2017). “Phylogenetic Classification of Bony Fishes Version 4”. BMC Evolutionary Biology 17 (1): 162. doi:10.1186/s12862-017-0958-3. PMC 5501477. PMID 28683774.
- ^ "Actinopterygii" (英語). Integrated Taxonomic Information System. 2006年4月3日閲覧。
- ^ R. Froese and D. Pauly: “FishBase” (February 2006). 5 July 2018時点のオリジナルよりアーカイブ。8 January 2020閲覧。
- ^ Van der Laan, Richard (2016). Family-group names of fossil fishes. doi:10.13140/RG.2.1.2130.1361
- ^ Xu, Guang-Hui (2021-01-09). “A new stem-neopterygian fish from the Middle Triassic (Anisian) of Yunnan, China, with a reassessment of the relationships of early neopterygian clades” (英語). Zoological Journal of the Linnean Society 191 (2): 375–394. doi:10.1093/zoolinnean/zlaa053. ISSN 0024-4082.
- ^ Nelson と ITISではトゲウオ目の亜目とされる。
参考文献
編集- [キャンベル11版] キャンベル生物学 原書11版. 丸善出版. (2018/3/20). ISBN 978-4621302767
- 原著:Lisa A. Urry; Michael L. Cain; Steven A. Wasserman; Peter V. Minorsky; Jane B. Reece; Neil A. Campbell (2016/10/29). Campbell Biology (11th Edition). Pearson. ISBN 978-0134093413
- 公益社団法人日本動物学会『動物学の百科事典』丸善出版、2018年9月28日。ISBN 978-4621303092。
- Joseph S. Nelson 『Fishes of the World Fourth Edition』 Wiley & Sons, Inc. 2006年 ISBN 0-471-25031-7
- 岩井保 『魚学入門』 恒星社厚生閣 2005年 ISBN 978-4-7699-1012-1
- 岡村収・尼岡邦夫監修 『日本の海水魚』 山と溪谷社 1997年 ISBN 4-635-09027-2