ハベリア
ハベリア(Habelia[4])は、約5億年前のカンブリア紀に生息した化石節足動物の一属。こぶのある体表と長い尾をもつ、カナダのバージェス頁岩で見つかったハベリア・オプタタ(Habelia optata)という1種のみによって知られる[5]。サンクタカリスなどと共に、ハベリア類という基盤的な鋏角類と思われるグループに分類される[2][3]。
| ハベリア | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 ハベリアの復元図
| ||||||||||||||||||
| 保全状況評価 | ||||||||||||||||||
| 絶滅(化石) | ||||||||||||||||||
| 地質時代 | ||||||||||||||||||
| 古生代カンブリア紀ウリューアン期 (約5億1,000万 - 5億500万年前)[1] | ||||||||||||||||||
| 分類 | ||||||||||||||||||
| ||||||||||||||||||
| 学名 | ||||||||||||||||||
| Habelia Walcott, 1912 [4] | ||||||||||||||||||
| タイプ種 | ||||||||||||||||||
| Habelia optata Walcott, 1912 [4] |
名称
編集学名「Habelia」はカナダブリティッシュコロンビア州のハベル山(Mount Habel)に由来する[4]。模式種(タイプ種)の種小名「optata」の由来は原記載(Walcott 1912)[4]に特記されていないが、ラテン語の「optatus」(願い・欲望)によると推測される[5]。
形態
編集-
![ハベリアの模式復元図(各項説明:[注釈 1])](//up.wiki.x.io/wikipedia/commons/thumb/a/a8/Habelia_diagram.jpg/866px-Habelia_diagram.jpg) ハベリアの模式復元図(各項説明:[注釈 1])
ハベリアの模式復元図(各項説明:[注釈 1])
![ハベリアの模式復元図(各項説明:[注釈 1])](/wiki/%E3%83%95%E3%82%A1%E3%82%A4%E3%83%AB:Habelia_diagram.jpg)
尾を除いて体長8.5mmから34mmまで及ぶ[2]。背面の外骨格は数多くのこぶが密生し、順に丸い頭部(前体)、12節のある胴部(後体)、および体長とほぼ同じ長さの尾節からなる[2]。
頭部
編集
頭部(cephalon, head, または前体 prosoma[2])は体長の約30%を占め、中央部分は丸く盛り上がり、中に大きな胃が収納される[2]。左右は三角形の平たい出っ張りで、一連の三角形の棘が縁の沿って生えている[2]。出っ張りの直前には1対の丸い切り込みがあり、1対の眼が附属する[2]。顕著な眼柄は見当たらず、複眼であったと考えられる[2]。この頭部は眼や口が由来する先節と直後7節の体節(第1-7体節)の融合でできた合体節で、真鋏角類における前体(先節+第1-6体節)と後体第1節(第7体節)に相同とされる[2][3]。
頭部腹面は順に三角形のハイポストーマ(hypostome)、左右2枚に分かれた上唇(labrum)、および7対の付属肢(関節肢)が並んでいる[2]。口は上唇と第2付属肢対の間で下向きに開き、常に第2付属肢対に覆われている[2]。
付属肢の中で最初の1対はかなり短小で、可動だが肢節数は不明、先端はおそらく爪がある[2]。この付属肢は鋏角に相同(第1体節由来)とされる[2][3]。
次の5対(第2-6対)は後方ほど発達した二叉型付属肢(biramous appendage)で、強大な顎基(gnathobase)、およびそれぞれ7節以上に分かれた歩脚型の外肢(exopod)と内肢(endopod)からなる[2]。顎基は前後重なりなって前に張り出し、頭部腹面のほとんどのスペースを占める[2]。顎基内側に並んだ歯は大小を繰り返し、左右相互に噛み合わせる[2]。歯は前の顎基ほど細く鋭く、後ろの顎基ほど鈍く頑丈に特化した[2]。内肢は7節からなり、第5-6肢節内側前端には9本以上の剛毛が集密し、先端の第7肢節は鉤爪状。関節は正面に折り曲げて、付け根は顎基の先端付近にあるため、内肢は常に先頭に集約されるように見える[2]。外肢は細長い7節以上からなり、3本の剛毛が各肢節の先端にもつ[2]。外肢のうち前4対は前方、第5対はやや離れて左右に突出する[2]。外肢の付け根の詳細は不明だが、顎基に関節しているようには見えず[2]、少なくとも前4対はそこから遊離したと考えられる[3]。この5対の付属肢は現生の真鋏角類における5対の歩脚型付属肢(触肢と残り4対の脚、第2-6体節由来)に相同とされる[2][3]。
第7対の付属肢は、直後の胴部前5節の付属肢に似たニ叉型付属肢である。ただし胴部のものに比べると、この付属肢の内肢は比較的に短く、外肢はもっと丸く周りの剛毛も比較的に多い[2]。正確には頭部と胴部の境目に付属しているが、常に下から頭部の腹面を覆いかぶさり、胴部付属肢のように対応の背板も見当たらないことから、機能的には頭部の付属肢とされる[2]。この付属肢は現生カブトガニ類の唇様肢とウミサソリの下層板に相同(第7体節由来)とされる[2][3]。
胴部
編集
胴部(trunk, または後体 opisthosoma[2])は12節でできた合体節で、各体節は12枚の発達した背板(tergite)によって表れる。各背板の表面は前半部が滑らかで、後半部が盛り上がって数多くこぶが密生していた[2]。各背板後半部の頂点に1対の突起があり、性的二形と思われる二形を示す。これは個体により前3対が角のように長く伸びる、もしくは前5対が少し大きくて後方ほど短くなる[2]。背板の左右は下方に曲がり、両端は後方ほど後ろ向きに湾曲し、前後の縁に鋭い棘が並んでいる[2]。この胴部は真鋏角類の後体第2-13節(第8-19体節)に相同とされる[2]。
体節数に応じて、計12対の付属肢が各胴節の腹面にある。前5対は発達した二叉型付属肢であり、内肢と外肢はそれぞれ円柱状の原節(basipod)の下側と外側に繋がり、原節基部は環状の節間膜を介して胴節に繋がる[2]。内肢は歩脚状で7節からなり、先端の肢節は鉤爪状で付け根はさらに2本の細い爪がある[2]。内肢のうち前3対はほぼ同じ長さだが、後2対は後方ほど若干長くなる[2]。外肢は2節でできたオール状で、第2節は楕円形に広く、外縁に分厚い剛毛が並んでいる[2]。
残り7対(第6-12対)の付属肢は単調な丸い外肢のみによって知られ、後方ほど小さく、内肢は退化的もしくはなかったと考えられる[2]。
このような付属肢の分化を基に、胴部の前5節を胸部(thorax、または中体 mesosoma)、後7節を「post-thorax」(または終体 metasoma)とも区別される[2]。
尾節
編集尾節(telson)は全長の半分以上を占めほど長く、途中の関節を介して前後2部に分かれている[2]。前半部の両筋は後ろ向きの棘が並び、基部は大きく膨らんで残りの部分は細長く、わずかに上向きに湾曲する[2]。基部腹側には「anal pouch」と呼ばれる袋状の突出部があり、肛門はその末端に開く[2]。後半部は前半部の1/3ほどの長さで真っ直ぐ伸ばし、3本の剛毛を末端にもつ[2]。
発見史と分類
編集ハベリア(ハベリア属 Habelia)はカナダブリティッシュコロンビア州のバージェス頁岩(Burgess Shale, カンブリア紀ウリューアン期、約5億1,000万 - 5億500万年前[1])で見つかった化石標本のみによって知られている[5][6]。本属の種として確実に認められるのは模式種(タイプ種)であるハベリア・オプタタ(Habelia optata)のみである。本属に含める可能性がある種 Habelia? brevicauda も知られているが[7][8][6]、Aria & Caron 2017 により本属から除外される[2]。
本属は20世紀初期から既に記載された化石節足動物である[4]が、前述の多くの特徴、特に頭部の付属肢とそれによって示される類縁関係は長らく解明されず、1世紀後の2017年の再記載で判明したものである[2]。
20世紀
編集

バージェス頁岩で見つかった多くの古生物(バージェス動物群)と同様、ハベリアは20世紀初期で古生物学者チャールズ・ウォルコット(Charles Doolittle Walcott)によって記載された。原記載(Walcott 1912)では、根拠は不確実でありながらも、ハベリアは光楯類に分類された[4]。数年後、ハベリアの頭部は少なくとも5対の付属肢があると他の研究によって確認された[9][10]。しかしこの見解は、古生物学者ハリー・ウィッティントン(Harry B. Whittington)の1981年の再記載(Whittington 1981)に否定され、ハベリアを節足動物における未詳化石(プロブレマティカ)の1つで[2]、頭部は1対の触角とおそらく2対の二叉型付属肢のみをもつと考えられた[8]。また長い間、ハベリアは頭部に眼をもたず、各胴節の背板に備わる突起は中央1本のみであるように復元された[5]。
2017年以降
編集-
 Aria & Caron 2017 に復元されたハベリア
Aria & Caron 2017 に復元されたハベリア -
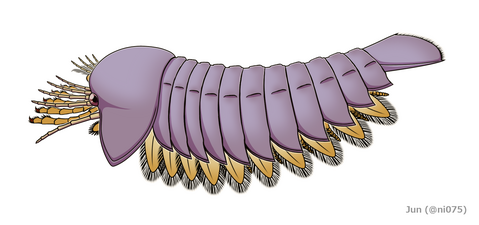


こうして数十年も再検討されなかったハベリアだが、Aria & Caron 2017 の再記載により、全身の復元像、特に頭部の付属肢構成が大きく書き替えられた[2]。頭部は眼をもち、強大な顎基をもつ5対の二叉型付属肢を備わり、前端は上唇や1対の小さな付属肢があり、背板の突起は対になるなど、今まで確認されていない構造を判明した[2]。中でも頭部左右の出っ張り・眼の位置・内肢の配置や構造が明らかにサンクタカリスに似て、近縁関係が示される。これにより、ハベリアを含くんだハベリア科とサンクタカリスなどを含くんだサンクタカリス科は、まとめて新設したハベリア目(Habeliida、またはハベリア類 Habeliid)に分類されるようになった[2]。他のハベリア類について、顎基など詳細の頭部構造はハベリアほどには明確でないが、知られる限り内肢はハベリアと同じく前方に集約するため、ハベリアのと同じ構造だと示唆される(内肢が顎基の先端に接続するからこそ可能の配置)。これにより、ハベリアに見られる頭部付属肢の構成は、ハベリア類全般の共有形質だと考えられる[2]。
| ||||||||||||||||||||||||||||||||||||||||||||||||
| 鋏角類におけるハベリアの系統的位置[3] |




ハベリアの特徴をはじめとして、ハベリア類の頭部付属肢の構成は、この類は基盤的な鋏角類であることを示唆する[2][3]。1対の短い付属肢と直後5対の同型な付属肢は、それぞれ鋏角類の前体における1対の鋏角と5対の歩脚型付属肢(触肢と脚)と対応しており、中でも発達した歩脚型の外肢は、基盤的な真鋏角類オファコルスとダイバステリウムにも見られる特徴である[2][3]。また、第7体節の付属肢が頭部の一部として機能することも、前体と融合し、その付属肢のように機能する一部の真鋏角類の第7体節(=後体第1節)付属肢(現生カブトガニ類の唇様肢・ウェインベルギナの第6対の歩脚型付属肢・オファコルスの鰭状の付属肢など[11])と共通している。これにより、真鋏角類の前体は、先節と次の6節だけでなく、通常では後体第1節とさる第7体節まで含むのが祖先形質であることも示唆される[2]。この説は、基盤的な真鋏角類とされ、第7体節の付属肢が前体のものとセットに特化したモリソニア[12]の発見によって更なる支持を得られている[3]。
付属肢の機能と生態
編集

ハベリアは基盤的な鋏角類とされるが、前体の付属肢はほとんど主に歩行に用いられ、後体の付属肢は多くが退化的な現生鋏角類[11]とは異なって、ハベリアの頭部(前体)付属肢は主に口器として用いられ、胴部(後体)の付属肢で歩行していたと考えられる[2]。このような付属肢の機能分化は現生の鋏角類らしからぬ、むしろ同様に節足動物だが別系統の大顎類(多足類・甲殻類・六脚類など)と収斂したとされる[2]。最も顕著な例はその強大な顎基に見られ、これは大顎類に特有の3対の口器である大顎(1対)と小顎(2対)のように、餌を粉砕・咀嚼できたと考えられる。ただし機能的分化が顕著な大顎(主に硬組織を粉砕する役を担う)と小顎(主に餌を細かく咀嚼する役を担う)とは異なって、ハベリアは5対の顎基があるにも関わらず全てがほぼ同形であった。これによりハベリアの顎基はまるで5対の大顎のように機能し、より単調で強大な粉砕力を発揮できたと考えられる[2]。他にも正面に突出した歩脚型外肢は(鋏角類にはなく、大顎類がつ)触角のように周りを探知し、第7対の付属肢は昆虫の下唇・一部の甲殻類の顎脚・ムカデの顎肢のように摂食を補助する付属肢であったと考えられる[2]。
これによりハベリアをも含んだハベリア類は、硬い外皮組織をもつ小動物を捕らえる小型捕食者であったと考えられる。ハベリア類はおそらくカンブリア紀における基盤的な鋏角類として、一部の基盤的な大顎類(例えばHymenocarina類)やArtiopoda類(例えばシドネイアなど)と共に、硬組織をもつ動物を狙う捕食者/腐肉食者のニッチ(生態的地位)を占める節足動物の一つであったと考えられる[2]。
脚注
編集注釈
編集- ^ a b
- am:胴部第1-5付属肢基部節間膜
- an:肛門
- ap:anal pouch
- bas:胴部付属肢原節
- ce1-5:頭部第2-6付属肢内肢
- cx1-5:頭部第2-6付属肢外肢
- das:肛門から押し出された内容物
- db:頭部第2-6付属肢内肢の剛毛
- dpex:胴部第1-5付属肢内肢第2節
- ds:胴部背側の突起
- dtp:尾節後半部
- e:眼
- e1-5:頭部第2-6付属肢内肢
- ex:頭部第7付属肢外肢
- g:顎基
- hyp:上唇
- ia:頭部第7付属肢
- it:消化管
- la:ハイポストーマ
- m:口
- oe:食道
- p1-7:頭部第2-6付属肢内肢第1-7肢節
- pex:胴部第6-12付属肢外肢
- ppex:胴部第1-5付属肢外肢第1節
- ptp:尾節前半部
- rap:頭部第1付属肢
- st:胃
- t1-5:胴部第1-5付属肢
- tel:尾節
- th:尾節基部
出典
編集- ^ a b c “How Old is the Burgess Shale” (英語). The Burgess Shale. 2023年1月13日閲覧。
- ^ a b c d e f g h i j k l m n o p q r s t u v w x y z aa ab ac ad ae af ag ah ai aj ak al am an ao ap aq ar as at au av aw ax ay az ba bb bc bd be bf Aria, Cédric; Caron, Jean-Bernard (2017-12). “Mandibulate convergence in an armoured Cambrian stem chelicerate” (英語). BMC Evolutionary Biology 17 (1): 261. doi:10.1186/s12862-017-1088-7. ISSN 1471-2148. PMC 5738823. PMID 29262772.
- ^ a b c d e f g h i j k Aria, Cédric; Caron, Jean-Bernard (2019-09). “A middle Cambrian arthropod with chelicerae and proto-book gills” (英語). Nature 573 (7775): 586–589. doi:10.1038/s41586-019-1525-4. ISSN 1476-4687.
- ^ a b c d e f g WALCOTT, C. D. 1912. Middle Cambrian Branchiopoda, Malacostraca, Trilobita and Merostomata. Smithsonian Miscellaneous Collections, 57: 145-228.
- ^ a b c d “Habelia optata” (英語). The Burgess Shale. 2023年1月13日閲覧。
- ^ a b “Habelia? brevicauda” (英語). The Burgess Shale. 2023年1月13日閲覧。
- ^ SIMONETTA, A. M. 1964. Osservazioni sugli artropodi non trilobiti della ‘Burgess Shale’ (Cambriano medio). III conributo. Monitore Zoologico Italiano, 72: 215-231.
- ^ a b Whittington, H. B. (1981-05-12). “Rare Arthropods from the Burgess Shale, Middle Cambrian, British Columbia” (英語). Philosophical Transactions of the Royal Society B: Biological Sciences 292 (1060): 329–357. doi:10.1098/rstb.1981.0033. ISSN 0962-8436.
- ^ Raymond, Percy Edward (1920). The appendages, anatomy and relationships of trilobites. New Haven, Conn.,: Connecticut Academy of Arts and Sciences. doi:10.5962/bhl.title.28256
- ^ Fedotov D. (1924). On the relations between the Crustacea, Trilobita, Merostomata and Arachnida. Bulletin de l'Académie des Sciences de Russie. 1924;383–408.
- ^ a b c Dunlop, Jason A.; Lamsdell, James C.. “Segmentation and tagmosis in Chelicerata” (英語). Arthropod Structure & Development 46 (3): 395–418. ISSN 1467-8039.
- ^ Gould, Stephen Jay; 渡辺, 政隆 (2000) (Japanese). ワンダフル・ライフ: バージェス頁岩と生物進化の物語. ISBN 978-4-15-050236-2. OCLC 676428996