ウミグモ綱
ウミグモ綱(ウミグモこう、学名: Pycnogonida)は、鋏角類に属する節足動物の分類群(綱)の1つ。構成種はウミグモ(海蜘蛛、ウミグモ類)と総称される[3]。胴体は小さく、体のほとんどが脚という独特な姿をした海棲動物である。名前と大まかな姿が似ているが、クモではない。
| ウミグモ | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
 様々なウミグモ[注釈 1]
| ||||||||||||
| 地質時代 | ||||||||||||
| 古生代カンブリア紀(約5億年前)[1][2] - 現世 | ||||||||||||
| 分類 | ||||||||||||
| ||||||||||||
| 学名 | ||||||||||||
| Pycnogonida Latreille, 1810 | ||||||||||||
| 和名 | ||||||||||||
| ウミグモ(海蜘蛛、ウミグモ類) 皆脚類 厚節類 脚体類 | ||||||||||||
| 英名 | ||||||||||||
| sea spider pycnogonid | ||||||||||||
| 目 | ||||||||||||
1,300種以上を知られ[4][5]、最古の化石記録はおよそ5億年前のカンブリア紀まで遡る[1][6]。現生種は皆脚目(かいきゃくもく、Pantopoda)のみからなる[7]。世界中の海域に広く分布するが、その生態と系統に関しては未解明な所が多い[8][9]。
名称
編集学名「Pycnogonida」はギリシア語の「pyknos」(厚い)と「gony」(膝)の合成語である[10]。唯一の現生目である皆脚目の学名「Pantopoda」はギリシア語「pantos」と「pous」の合成語で、発達した脚が体の大部分を占める姿に因んで名付けられた[10]。
英語は学名に因んだ「pycnogonid」のほか、海棲でクモ(spider)に似た姿から一般に「sea spider」と呼ばれる[10]。和名も同じ意味で「ウミグモ」(海蜘蛛、ウミグモ類)といい、他にも皆脚類(かいきゃくるい、Pantopoda)・厚節類(こうせつるい、Pycnogonida)・脚体類(きゃくたいるい、Podosomata)などと総称される[3][11]。古くは夢虫(ユメムシ)とも呼ばれ、21世紀ではユメムシ科にその名が残っている[12]。
形態
編集_003.jpeg)



小さな胴体に対して8本(稀に10本と12本[注釈 2])の脚は極端に発達し、脚を束にしただけのような姿をした種類が多いが、ヨロイウミグモ科のように丈夫な体型をもつものもある[5]。左右の脚を広げた大きさは微小な数 mmから大型の数十 cmまで達し[10][5][13]、多くの種類は脚を広げて5 mmから3-4 cmに及ぶ[10]。知られる中で最大種は体長8.5 cm・脚を広げて70 cmに達するベニオオウミグモ、最小種は体長1 mm未満・脚を広げて2 mm の Austrodecus palauense である[10]。体色は色薄いものから Anoplodactylus evansi のように鮮やかなものまで様々である[5]。
体節の構成
編集_002.jpeg)
体の前方は先節とそれぞれ鋏肢・触肢・担卵肢・第1脚をもつ4節の体節の融合でできた合体節で、それ以降はそれぞれ1対の脚をもつ3節(稀に4-5節)の体節と付属肢をもたない体節が続く[14]。いずれも円筒状で、外骨格は他の節足動物で一般に見られる明確な上下区別(背板 tergite と腹板 sternite)はない[14]。
なお、これらの部位に対する区分は昔今の文献によって異り、以下の例が挙げられる[15][16][17][18](5-6対の脚をもつ希少例における5-6番目の脚と脚4対の通常例の体節対応関係は2017年現在では未検証[14]のため、以下の表は通常の脚4対の通常例のみについて扱う)。
| 体節の番目 | 先節(0) | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8-n |
|---|---|---|---|---|---|---|---|---|---|
| 付属肢(関節肢) | - | 鋏肢 | 触肢 | 担卵肢 | 第1脚 | 第2脚 | 第3脚 | 第4脚 | - |
| 外見上の節の番目 | 1 | " | " | " | " | 2 | 3 | 4 | 5-n |
| 1:古典的体系[15] | △ | △ | △ | △ | ◎ | ◎ | ◎ | ◎ | * |
| 2:主流な体系[15][16] | △ | △ | △ | △ | △ | ◎ | ◎ | ◎ | * |
| 3:前体・後体体系(1)[16] | △ | △ | △ | △ | △ | △ | △ | * | * |
| 4:前体・後体体系(2)[16][17][10] | △ | △ | △ | △ | △ | △ | △ | △ | * |
- 鋏肢・触肢・担卵肢をもつ部分を「頭部」、脚をもつ部分を「胴部/胸部」、後端の部分を「腹部」と扱う[15][19]。
- 鋏肢・触肢・担卵肢・第1脚をもつ部分を「頭部」、残りの脚をもつ部分を「胴部/胸部」、後端の部分を「腹部」(もしくは「trunk end」[20])と扱う[14][16][17]。
- 鋏肢・触肢・担卵肢・第1-3脚をもつ部分を「前体」、第4脚(第7体節)以降の部分を「後体」と扱う(真鋏角類の体制に従う)[16]。
- 鋏肢・触肢・担卵肢・脚をもつ全ての部分を「前体」、後端の部分を「後体」と扱う(鋏角類の第7体節を前体に含める体制に従う)[16][17][21]。
特に注目される部分として、古典の体系(例1)に「頭部」(head)とされた部分(先節+第1‐3体節)は、直後の第1脚をもつ第4体節と完全に融合して合体節をなしている(化石種 Palaeomarachne granulata のみその境目らしき痕跡が見られる[22])。特に現生の真節足動物の先頭の合体節(頭部融合節)は先節を除いて4節以上含めるため、形態学上での比較のため、20世紀後期以降の文献記載ではこれらの4節を含んだ合体節を「頭部」(cephalon または cephalosoma[15])と扱うのが主流となっている(例2)[15][16][14]。
本項目では、主流な体系(例2)に基づいてウミグモの特徴を記述する。

頭部
編集頭部(cephalon、cephalosoma[15])は先節と直後4節の体節(第1-4体節)からなり、全てが融合して1つの合体節をなしている[23][15][14]。なお、この頭部は頭らしい形とは言いがたく、第1脚をもつ体節(第4体節、古典の体系における「第1胸節」)をも含め、その構造も胴節と同形である[14]。この頭部は他の一部の鋏角類(ヒヨケムシ、コヨリムシなど)における分節した前体の前半部に対応とされる[14]。
頭部の背面中央は眼丘(ocular tubercle)が盛り上がり[19]、通常はここで2対の単眼をもつ。これらの単眼は全て中眼(median eye)由来で、側眼(lateral eye, 祖先形質として複眼)由来のものは存在しない[24]。なお、1対や無眼まで退化し[25]、眼の代わりに感覚器として機能する突起 (lateral sense organ) をもつ種類もある[26]。
頭部の前面はよく発達した吻(proboscis)が突出し、形は円筒状から徳利のように丸みを帯びたものがある。この吻は通常では三放射対称の構造で、先端は3つの可動の構造体(lip, jaw)に囲まれた逆三角形の口が開いている[27][8]。ただしスイクチウミグモ科の場合、口は吻の先端腹側に開いた縦長いスリット状である[4]。一部の種類では吻の付け根に関節があるため、吻を体の腹側に折り畳める[4]。この吻は節足動物の中でウミグモに特有で、その由来や他の節足動物の部分との対応関係(相同性)は不明確である[14]。他の節足動物の口器に見られる上唇や頭楯/口上板/ハイポストーマに類する構造は存在しない[14]。
頭部は基本として計4対の付属肢(関節肢)をもち、順に先節直後の第1-4体節に由来する。鋏肢・触肢・担卵肢という古典的体系における3対の「頭部付属肢」、および第1脚からなる。ただし前3対の「頭部付属肢」は、分類群や雌雄によって成長する度に退化消失した場合がある[19][8][9]。これらの付属肢の有無と構造は伝統的に科の同定形質として重要視されていた[19][5]。ただし2010年代以降、この識別方法は一部の科においては系統関係を反映できず、疑問視されつつある(後述参照)[28]。

![鋏肢と触肢をもたない Pycnogonum littorale(ヨロイウミグモ科)、この類では雌が担卵肢をも欠く[9]。](/wiki/%E3%83%95%E3%82%A1%E3%82%A4%E3%83%AB:Pycnogonum_littorale_(YPM_IZ_030249).jpeg)
.jpeg)
鋏肢
編集
A, B: Ascorhynchus abyssi、C, D: Ammothea hispida
-
 Cordylochele longicollis の頭部、吻と鋏肢
Cordylochele longicollis の頭部、吻と鋏肢 -
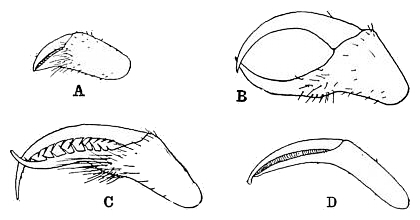
-
 Decolopoda australis の4節の鋏肢
Decolopoda australis の4節の鋏肢



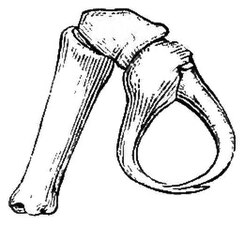
吻の直後に鋏肢(きょうし、chelifore, cheliphore[15])があり、これは第1体節由来の付属肢で、他の鋏角類の鋏角(きょうかく、chelicera)に該当する[14]。鋏肢は種類によって背側(ホソウミグモ科など)もしくは左右に寄せる方向(カニノテウミグモ科、ユメムシ科)で備わっている。後者の場合、鋏肢の可動域は広く、往々にして頑強で、獲物を握りつぶす役割に適している[8]。通常は3節で先端2節ははさみをなしているが、柄部(scape)に該当する第1肢節が更に2-3節に細分され、計4-5節になった種類もある[注釈 3][15][20][14]。なお、鋏肢が成長に連れて短縮し、果ては成体で完全に消失した種類もある(ヨロイウミグモ科、オオウミグモ科など)[8]。
触肢
編集鋏肢の次、頭部の左右に1対の触肢(しょくし、palp)がある。これはクモガタ類の触肢(pedipalp)と同じ第2体節由来の付属肢である[14]。感覚や食物の把握などに用いられ[8]、肢節数・発達具合・および触肢そのものの有無は分類群(一部の群では雌雄)によって異なる[19]。従来では4-10節の肢節をもつとされていた[3][19]が、最多の10節をもつと思われるオオウミグモ属(Colossendeis)の触肢における「第1節」とされた付け根の膨らみは、真の肢節ではなく、単に頭部外骨格の延長部(lateral process)だと後に判明した。これによれば、ウミグモの触肢の肢節数は最多9節となる[28]。先端は通常では単調であるが、一部の化石種では1本の鉤爪がある[18]。
担卵肢
編集
A: Phoxichilus spinosus、B: Phoxichilidium femorattum、C: Anoplodactylus petiolatus、D: Colossendeis proboscideus
-
 Anoplodactylus lentus の担卵肢(左)と第4脚(右)
Anoplodactylus lentus の担卵肢(左)と第4脚(右)
_002.jpeg)
触肢と第1脚の間には、担卵肢(たんらんし、oviger, ovigerous leg[9])というウミグモに特有[19]の細長い付属肢がある。発生学と解剖学的構造は触肢に似ている[注釈 4][17]が、これはクモガタ類の第1脚に相同(第3体節由来)の「特化した脚」だと考えられる[17][14]。普段は体の下に折り曲げ、先端は身繕い用の棘状突起が並ぶ[19]。雄は卵塊をこの付属肢につけて保護する[9]。肢節数は分類群によって4-10節からなる[3]。なお、第1肢節の直前に連結する外骨格の膨らみも肢節とし、肢節数を最多11節とする見解もあるが、それは単なる頭部外骨格の延長部だと後に判明したため、否定的とされる[28]。通常は雌雄とも担卵肢をもつが、ヨロイウミグモ科とホソウミグモ科などでは雌が担卵肢を欠く[9]。ヨロイウミグモ属(Pycnogonum)の Nulloviger 亜属では、雌雄とも担卵肢を欠く[9]。
胴部
編集.jpeg)
胴部(trunk, 胸部 thorax ともいう[15])は幅が狭く、脚の太さとほぼ変わらない。基本として3節の胴節(trunk somites, 第5-7体節)を含め、5-6対の脚をもつ種類では脚の数に応じて胴節が4-5節まで追加される[29]。各胴節は通常では明瞭な関節に分かれているが、直前の頭部をも含んで全てが融合した例もある[25]。各胴節の左右からやや延長した接脚突起(lateral processes)[19]が脚に連結する。この胴部は真鋏角類における前体最終2節と後体第1節に対応とされる[14]。
脚
編集
1: Chaetonymphon hirtum、2: Nymphon stroemi、3: Nymphon brevirostre、4: Ammothea echinata、5: Ascorhynchus abyssi

-
 Endeis spinosa の第1脚
Endeis spinosa の第1脚 -
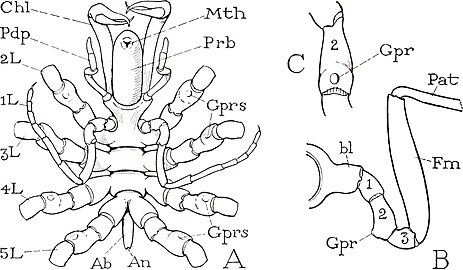 Chaetonymphon spinosum の生殖孔(Gpr)
Chaetonymphon spinosum の生殖孔(Gpr)
_001.jpeg)

脚(歩脚、歩行肢、legs, walking legs, ambulatory legs)は体長の倍以上に発達で、体の大部分がこの付属肢からなる[10]。通常は4対で、頭部第4節から第3胴節(第4-7体節)にかけて備わっているが、後ろから1-2対の脚を増やされ、計5対や6対をもつ種類も稀わずかにある[注釈 2][30][10][29][14]。
ごく一部の化石種(後述)を除いて、脚は全て同形な歩脚型で、先端の爪を除いて見かけ上8節の肢節から構成される[19]。第1-3節は短く、そのうち第2肢節は生殖口(gonopore)をもち、通常は腹面に開く[注釈 5][9]。パレオイソプスやハリエステスなどの古生代の種類は、第1節は更に数個の幅狭いリング状の構造 (annulation) に細分されている[15]。第4-6節は長く発達し、先端の第7-8節は短い。第8節先端に1本の爪(主爪、main claw)があり、その付け根の上方が更に1対の短い副爪(auxiliary claws)をもつ場合もある[31][19]。接脚突起と第1肢節の関節、および第2から第6節までの4つの関節は上下方向に、第1節と第2節の関節は前後方向に動けるようになり、それぞれ1対の筋肉(伸筋と屈筋)に操られている[10]。一方、第6-7節の関節は腹面1つの屈筋のみをもち、第7-8節の関節は筋肉に繋がれていない[10]。
なお、化石群には上述の特徴に当てはまらない例外がある。例えばパレオイソプスの脚は前方ほど長いオール状の遊泳脚で第1脚が第2-4脚より1節少なく、パレオパントプスの脚は9節をもつ[15][26]。
脚の肢節の呼称と解釈は文献記載によって異なる場合があり、次の表の通りに挙げられる。1番目の体系は常用されるものだが、転節がない・趾節という独自の呼称を使われるなど、通常の節足動物のものとは大きく異なる[10]。残り2つの体系は相同性を踏まえて他の鋏角類の脚と統一されたもので、転節と膝節があり、そのうち3番目の体系(腿節2節説)は筋肉の構造により有力視されている[32]。
肢節の番目 体系
|
1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 爪/9 |
|---|---|---|---|---|---|---|---|---|---|
| 通常(転節・膝節なし)[19][10] | 第1基節 coxa 1 | 第2基節 coxa 2 | 第3基節 coxa 3 | 腿節 femur | 第1脛節 tibia 1 | 第2脛節 tibia 2 | 跗節 tarsus | 趾節 propodus | (趾節 propodus の一部) |
| 転節・膝節あり、転節2節[15] | 基節 coxa | 第1転節 trochanter 1 | 第2転節 trochanter 2 | 腿節 femur | 膝節 patella | 脛節 tibia | (跗節 tarsus の一部) | (跗節 tarsus の一部) | (跗節 tarsus の一部) |
| 転節・膝節あり、腿節2節[32] | 基節 coxa | 転節 trochanter | 第1腿節 femur 1 / 前腿節 prefemur / basifemur | 第2腿節 femur 2 / 後腿節 postfemur / telofemur | 膝節 patella | 脛節 tibia | (跗節 tarsus の一部) | (跗節 tarsus の一部) | 前跗節 pretarsus / apotele |
| 転節なし・膝節あり[18] | 第1基節 coxa 1 | 第2基節 coxa 2 | 第3基節 coxa 3 | 腿節 femur | 膝節 patella | 脛節 tibia | (跗節 tarsus の一部) | (跗節 tarsus の一部) | 前跗節 pretarsus / apotele |
また、脚を4対もつという点からクモガタ類の鋏角類を彷彿とされるが、担卵肢の存在によってこれらの脚の対応関係(相同性)はクモガタ類のものとは1節ずれていて、順番は相同でなく、特に第4脚は、他の現生鋏角類に脚を持たない後体第1節(第7体節)に対応するとされる(次の表および後述の議論も参照)[14]。
体節 分類群
|
1 | 2 | 3 | 4 | 5 | 6 | 7 |
|---|---|---|---|---|---|---|---|
| 真鋏角類 | 鋏角 | 触肢 | 第1脚 | 第2脚 | 第3脚 | 第4脚 | 唇様肢(カブトガニ)/下層板(ウミサソリなど)/退化(クモガタ類) |
| ウミグモ類 | 鋏肢 | 触肢 | 担卵肢 | 第1脚 | 第2脚 | 第3脚 | 第4脚 |
腹部
編集腹部(abdomen, trunk end[20])は皆脚目の種類ではごく小さく、体節構造が見られない粒のような部分であり、末端に肛門がある。ただし一部の化石種では、腹部は比較的に発達し、複数の体節と1本の尾節(telson)をもっていた[14][15][33]。この腹部は真鋏角類における後体第2節以降の部分に対応とされる[14]。
内部構造と生理学
編集


吻は筋肉質で、内側(咽頭)はフィルターらしき繊毛構造が並んでいる[27][34][8]が、確実の機能は不明[8]。消化管(中腸)は枝分かれして脚と鋏肢に入り込んで、多くの種類ではこれらの付属肢の先端近くまで伸びている[8][35][36]。触肢と担卵肢ではこのような消化管の枝を欠く[17]。生殖腺は体腔の背側に格納され、これも枝分かれして脚に入り込んでいる[9]。雌の場合、卵巣は脚の腿節に格納される[9]。
他の節足動物と同様にはしご形の中枢神経系をもち、3つの脳神経節(それぞれ先節と第1-2体節由来の前大脳・中大脳・後大脳[37])のうち前大脳と中大脳は食道孔の前に癒合し、その直後の後大脳は担卵肢(第3体節)の神経節と癒合する[36]。それ以降の腹神経索は脚に対応する神経節が梯子状に並んでおり、後方の神経節が前の胴節に移行した場合がある[36]。真鋏角類ほどの癒合(中枢神経系全体が高度に癒合し、リング状の synganglion をなしている)はないが、分類群により脚の神経節が前後に密着・担卵肢と第1脚の神経節が癒合する場合もある[36]。最終胴節の神経節の直後にあり、腹部由来と思われる退化的な神経節は幼生の段階で一時的に見られ、発育が進む度に胴節の神経節に吸収される[38]。
ウミグモの多くの臓器は脚に格納されており、多くの生理的活動も脚が荷う。鰓など独立した呼吸器官はなく、脚の多孔質な外骨格を通じて直接にガス交換を行う(皮膚呼吸)[39]。胴部背側にある心臓は貧弱で、その脈動で流される血リンパは脚の基部にしか届かず[35]、果ては心臓が退化消失した分類群もある[36]。その代わりに、脚にある大部分の血リンパと酸素を胴部に行き渡して全身の血液循環を果たすには、脚に入り込んだ消化管の枝が波打つるように、順調よく伸縮して血リンパを流動させるという独特な方法で行う[35][40]。ウミグモのこのような脚と消化管の枝は、それぞれ鰓と心臓のような機能をしているとも形容される[35]。また、生殖口と生殖腺は脚にあるため、産卵と放精も脚を通じて行う[9]。
雌雄
編集一般的な性的二形として、雄は雌に比べてやや小型となる[9]。ヨロイウミグモ属とホソウミグモ科などの場合、雌は担卵肢を欠く[9]。
明確に雌雄を区別できる特徴は生殖口にある。雄の生殖口は微小な穴で、蓋状構造を欠く場合もあるのに対して、雌の生殖口は大きく、往々にして蓋状構造をもつ[9]。生殖口の数も往々にして雌雄が異なる。雄は属によって前の数対の脚が生殖口を欠く場合があるが、雌は一部の例外[注釈 6]を除いて全ての脚に生殖口をもつ[9]。また、腿節に格納されるセメント腺 (cement gland) は雄に特有の器官である[9]。
生態
編集

寒帯から熱帯の海域、潮間帯から数千メートルの深海まで広く分布する海棲動物である[3]。南極と亜南極の海域に生息するものは260種以上が知られている[41][28]。
運動が緩慢な底生動物であり、海底の岩・海藻・サンゴ礁などにしがみつき、もしくは堆積物の表面をゆっくりと歩行をし、前者は脚が丈夫な種類、後者は脚が細長い種類に多い[10]。時には脚を上下に連動させて、水中を泳ぐこともある[10]。軟体動物や刺胞動物に寄生するもの、自由生活のものなどがある[42][8][7]。トックリウミグモ属(Ascorhynchus)[43]は普段が脚を平たく畳んで堆積物に潜む[44]。
いくつかの種類は、刺胞動物・カイメン・コケムシなどの付着生物に口吻を刺し込んで体液を吸収することが知られているが、他のものについては食性がほぼ不明[8]。一部の種類は、口が金魚のように開閉する様子が観察されており、これは濾過摂食に関与する動きではないかと考えられる[8]。また、ヤマトトックリウミグモについては、2020年に鳥羽水族館で飼育されている個体がイソギンチャクを捕食する様子が確認された[45]。古い文献では「吻で二枚貝を殻を突き抜いて捕食する」「クジラに寄生する」という文章記述もあったが、どれも懐疑的で、特に後者は単にクジラジラミ(クジラに寄生し、ヨロイウミグモ科に似る姿をした甲殻類)の見間違いである[8]。
天敵と自衛手段に関してはほぼ不明である。魚類やカニなどの肉食動物に襲われる様子は目撃されているが、捕食に至らず、すぐ放されている[46]。少なくともヨロイウミグモ科の1種 Pycnogonum litorale は、高い濃度の脱皮ホルモンで捕食者に嫌がれることと[47]、幼体が欠損した後半身まで再生できるほど強力な再生能力をもつことが判明した[48]。体表はときおりにエボシガイなどの付着生物がくっついて、場合によってはウミグモの呼吸と運動に悪影響を与える[49][46]。抱卵中の雄は通常の個体に比べて捕食者に狙われやすく、付着生物にくっつけられやすいことを示唆する研究がある[50]。また、一部の種類は Dickdellia 属の寄生性腹足類に宿主とされる[51][52]。
繁殖と発育
編集-
 対になるウミグモ
対になるウミグモ -
 抱卵中のウミグモ
抱卵中のウミグモ -
 卵嚢を担卵肢にもつ Nymphon molleri
卵嚢を担卵肢にもつ Nymphon molleri



_004.jpeg)
繁殖は体外受精を通じて行う[9]。受精のたびに、雄は雌に抱きつき、お互いの生殖口が近づける姿勢をとる。この姿勢は、通常では雄が雌の背中に乗るが、Anoplodactylus 属のように、雄が裏返して雌の腹側に張り付くなどやや異なった例もある[9]。雌は脚の生殖口から卵を産み、雄はそれを受け取って、セメント腺から分泌した粘液で卵嚢を担卵肢に粘りついて世話にする[9][50]。
他の繁殖行動はほぼ不明だが、一部の種類からは、雄が雌を追いかけ、雌同士の雄を奪い合うための闘争行為が確認される[9]。
発生は甲殻類のノープリウス幼生を彷彿とさせる[53]。多くの場合はプロトニンフォン幼生(プロトニンフォン、protonymphon)という特殊な段階で孵化し[7][54]、体は丸く、鋏肢・触肢・担卵肢という前3対の付属肢のみをもち、触肢と担卵肢は成体らしからぬ爪のような形をした[53]。これは脱皮を経て徐々に成体に近い姿に変態しては、脚をもつ体節を順に後ろから追加される[55]。なお、脚をもつプロトニンフォンや成体の姿に近い段階など、比較的発育が進んだ姿で孵化する種類もある[53]。
化石
編集現生種の普遍性に反して、化石ウミグモ類の発見は非常に稀である[56]。特に中生代以降の化石記録は、長い間に欠けていた[6]。ただし現生種に比べると、古生代のウミグモ類は体制的に多様で、これは主に腹部の構造で明瞭に表れる[14]。
古生代
編集

ほとんどの化石記録は古生代によるものである。既知最古の化石標本は、約5億年前のカンブリア紀後期まで遡るカンブロピクノゴン (Cambropycnogon) の幼生化石である。このプロトニンフォン幼生は、付属肢の基部内側に突起(顎基)をもち、体の尾端に1対の長い付属体があるなど、現生群に見当たらない形質をもつ[1]。この化石のウミグモ類としての形質を疑う研究もある[57]が、否定的と評価される[58][56]。
Cambropycnogon に次いては、約4億5,000万年前のオルドビス紀後期に生息したパレオマラクネ (Palaeomarachne) がある。この種は断片的な本体部分しか発見されていないが、頭部は他のウミグモ類に見当たらない体節の境目的な痕跡をもつ[22]。
シルル紀前期(約4億2,500万年前)のハリエステス(Haliestes)[20]は、体長数 mmの小型種であるにもかかわらず、保存状態がかなり良好な化石標本によって知られ、完全な成体化石をもつウミグモの中では既知最古の種となる。本種は一見して現生群に似ているが、脚基部の環形構造や腹部の分節など、後述のデボン紀の種類に似た特異な性質も見られる[59]。
デボン紀
編集


デボン紀前期(およそ4億年前)の堆積累層フンスリュック粘板岩(Hunsrück Slate)からは、比較的に多くの化石ウミグモ類が見つかり、パレオイソプス (Palaeoisopus)、パレオパントプス(Palaeopantopus)、パレオテア (Palaeothea)、フラジェロパントプス(Flagellopantopus)、ペンタパントプス (Pentapantopus) の5属が知られている[15][26]。現存種とはかけ離れた形質をもつ化石種は多くがここで知られており、ウミグモ類の系統における体制の多様性を大きく拡張した[14][33]。
パレオイソプスは特に代表的で、多くの化石標本が発見される[2]。この種は、4節の長い腹部・剣状の尾節・遊泳生活に適したへら状の脚・特化した第1脚・5節の鋏肢など多くの特異な形質をもつ[15]。パレオパントプス とフラジェロパントプスは短いながらも数節に分かれた腹部をもち[15]、特に後者は胴長の2.5倍に達する鞭状の長い尾節をもっていた[60]。また、この2属の脚の第4節が不動な2節に細分されていたのも特徴的である[26]。
中生代
編集
2023年現在、中生代のウミグモは未だに下記のジュラ紀の属のみによって知られ、直前の化石記録(デボン紀)に比べても2億5000万年ほどのギャップを開いている[2][18]。古生代のものとは異なり、中生代のウミグモは知られる限り全て皆脚目に属している[18]。
中生代ウミグモの化石記録として、かつてペンタパレオピクノン (Pentapalaeopycnon) とピクノゴニテス (Pycnogonites) という2属はあったが、いずれも単に十脚目甲殻類のフィロソーマ幼生の見間違いだと後に判明した[1]。こうして実際が長い間に欠けた中生代の化石記録は、2007年に記載されたLa Voulte Lagerstätte産のジュラ紀(1億6000万年前)ウミグモ類が本格的に初の記載となる[6]。この堆積累層にはパレオピクノゴニデス (Palaeopycnogonides)、コロッソパントプス (Colossopantopodus)、パレオエンデイス (Palaeoendeis) の3属が知られ、Palaeopycnogonidesは独自の科 (Palaeopycnogonididae) に属するが、Colossopantopodus はオオウミグモ科に、 Palaeoendeisはミドリウミグモ科に分類される[18]。前述したものより少し後(ジュラ紀後期、1億5000万年前)なゾルンホーフェン石灰岩には、2種目のコロッソパントプス (Colossopantopodus nanus) と、現生のEurycyde属に属する可能性がある1種が知られている[2]。
頭部付属肢の対応関係
編集
体節 分類群
|
先節 (前大脳) |
1[注釈 7] (中大脳) |
2[注釈 7] (後大脳) |
3 | 4 | 5 |
|---|---|---|---|---|---|---|
| ウミグモ類(最も有力視される説) | ? | 鋏肢 | 触肢 | 担卵肢 | 第1脚 | 第2脚 |
| ウミグモ類(触肢・担卵肢同一体節由来説) | ? | 鋏肢 | 触肢+担卵肢 | 第1脚 | 第2脚 | 第3脚 |
| ウミグモ類(鋏肢前大脳性/先節由来説) | 鋏肢 | 触肢 | 担卵肢 | 第1脚 | 第2脚 | 第3脚 |
| 真鋏角類 | 上唇 | 鋏角 | 触肢 | 第1脚 | 第2脚 | 第3脚 |
| 大顎類 | 上唇 | 第1触角 | 第2触角/(退化) | 大顎 | 第1小顎 | 第2小顎/下唇 |
| ラディオドンタ類 (前部付属肢前大脳性説) |
前部付属肢 | 鰭 | 鰭 | 鰭 | 鰭 | 鰭 |
ウミグモの鋏肢は、通常では真鋏角類の鋏角に相同で、すなわち第1体節由来(中大脳性)の付属肢と見なされる。しかしウミグモの鋏肢神経は、一見して脳の先頭に対応するようにも見える。この特徴に基づいて、ウミグモの鋏肢は真鋏角類の鋏角に非相同で、むしろ先節由来/前大脳性(ラディオドンタ類などの前部付属肢に相同の可能性がある)ではないかという異説は一時的に提唱された[61]。しかしこの見解は、公表されるあとで多くの研究に否定的と評価される。ホメオティック遺伝子発現[62][17]や神経発生と神経解剖学的再検証[37]は、いずれも通説の鋏肢第1体節由来/中大脳性説を強く支持し、鋏肢神経に対応する脳の先頭は前大脳ではなく、前方に曲がった中大脳だと示される[37]。
また、担卵肢の対応関係も議論の的とされていた。これは真鋏角類の第1脚(第3体節に対応)に相同というのが通説だが、別の異説が2つある。1つは、担卵肢と触肢の解剖学と発生学上の共通点[注釈 4]や、一部の群(オオウミグモ科など)での基部がお互いに密着するなどの形質を根拠とし、両者は同一体節由来で、担卵肢は「重複した触肢」という説である[17]。もう1つの説は、前述の触肢との同一体節由来を否定しつつも、上述の鋏肢先節由来/前大脳性説を踏まえて、担卵肢を真鋏角類の触肢に相同と見なしている[61][17]。しかしホメオティック遺伝子発現は、通説(触肢とは別の第3体節由来)を支持し、これらの異説を否定している[17]。
分類
編集系統位置
編集
| |||||||||||||||||||||
| ウミグモの最も広く認められる系統的位置(鋏角類説) |
| |||||||||||||||||||||
| 幹性類説 |
古くは海産のクモガタ類や極端に特化した甲殻類とも解釈されていたが、いずれの見解も後に否定的とされる[63]。
通常、ウミグモ類は節足動物のうち鋏角類(鋏角亜門)の1綱として分類されている。しかし2000年代初期での分子系統解析により、ウミグモ類の単系統性は強く支持されるものの、他の鋏角類との類縁関係は一時的に大きく動揺された[63]。
2000年代以降の分子系統解析ではさまざまな結果が出て、中でもウミグモ類は他の現生鋏角類(真鋏角類)に類縁でなく、残り全ての現生節足動物(幹性類 Cormogonida)の姉妹群になるという異説が特に注目される(幹性類説)[64][65]。この場合、ウミグモは鋏角類に含まれない、もしくは鋏角類は解体され、真鋏角類とウミグモはそれぞれ独立の亜門(真鋏角亜門、ウミグモ亜門/皆脚亜門[11])に昇格される[66]。しかし、様々な解析結果のなかでも、ウミグモと他の鋏角類(真鋏角類)が姉妹群になり、すなわち鋏角類に含める系統関係の方が多数派であり[67][17][68][69][70][71][5]、特に2010年代以降の系統解析から再び支持が得られつつある[69][70][71][72][73]。他にもクモガタ類に含まれ、そのうちダニ類やコヨリムシ類に近い系統関係が与えられた[74][75][76]が、これらは長枝誘引がもたらす誤推定として疑問視される[77]。また、ウミグモ類の形態は特異のため、基盤的な鋏角類とされる化石節足動物(ハベリア・サンクタカリス・モリソニアなど)と比較しにくく、関係性は難解である[78]。
鋏角類説と幹性類説は、いずれも分子系統学以外に形態学と発生学的根拠が挙げられる。前者は、第1対の付属肢は基本としてはさみ型の鋏角/鋏肢、および全面的な付属肢構成と相同性に支持される[17][14]。後者は、上唇の欠如・付属肢にある複数対の生殖口などという、ウミグモ類以外の節足動物に見当たらない形質が挙げられる。しかし後者については、幹性類説を支持できる節足動物の祖先形質か、それとも単にウミグモの派生形質なのかどうかは不明確である[17]。また、前述の鋏肢先節由来/前大脳性説も幹性類説を支持する証拠と見なされた[61]が、その対応関係は後に多くの再検討に否定されるようになった[62][17][37](前述参照)。
下位分類
編集2015年まででは、79属1385種のウミグモが記載される[5]。唯一の現生目である皆脚目(Pantopoda)と、それぞれ1種のみからなる3つの絶滅目に分かれ、ほかにも分類未解明の化石種がいくつかある[79]。これらの群のお互いの系統関係については、文献によって意見が分かれ、定説はないが、少なくともパレオイソプスと Flagellopantopus は基盤的なウミグモ類である説は比較的に広く認められる[56]。
- ウミグモ綱 Pycnogonida
- †(属)Cambropycnogon
- †(属)Palaeomarachne
- †(属)Flagellopantopus
- †(属)Palaeothea
- †(属)Pentapantopus
- †古皆脚目[80](ムカシウミグモ目[81][82])Palaeopantopoda
- ムカシウミグモ科[82] Palaeopantopodidae
- †パレオイソプス目 Palaeoisopoda
- パレオイソプス科(ウミユリヤドリグモ科[82]) Palaeoisopodidae
- †(目)Nectopantopoda[85]
- 皆脚目[7](真皆脚目[80]、ウミグモ目[81][82]) Pantopoda
- (後述参照)
皆脚目
編集
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Ballesteros et al. 2020 に基づいた皆脚目の内部系統関係[87]。 |

.jpeg)
_(14206708079).jpg)
_002.jpeg)
.jpeg)
_002.jpeg)

_002.jpeg)

皆脚目(Pantopoda)の現生種は、昔今の見解によって9-11科に区別される。その内部系統関係は議論的であり、分子系統解析の進展により再編成をなされつつある[7]。特に外部形態に基づいた従来の9科の一部の単系統性と同定形質の有効性は、徐々に疑問視される向きがある[5]。例えば、かつては類縁関係を推測できると考えられた頭部付属肢の退化具合は、分子系統解析によると、これは多くが複数の群から独自に獲得した特徴(収斂進化)だと示唆される[28]。
スイクチウミグモ科・ヨロイウミグモ科・オオウミグモ科の3群の他の群より早期に分岐した系統関係は広く認められ[4][88]、2020年ではそのうちスイクチウミグモ科は最も基盤的なウミグモで、残りの2群はイボウミグモ科と単系統群になる系統関係が有力視される[87]。カニノテウミグモ科は、ユメムシ科を除いた側系統群だとされる[4][88][87]。
2018年現在、現生種の2亜目6上科11科の分類体系は以下の通り[89][7]。なおこの分類体系は、上述の分子系統解析に示唆される系統関係を適切に反映できない問題がある。
- 皆脚目 Pantopoda
- (亜目)Eupantopodida
- (上科)Ascorhynchoidea
- (上科)Colossendeidoidea
- (上科)Nymphonoidea
- (上科)Phoxichilidoidea
- (上科)Pycnogonoidea
- (上科)Rhynchothoracoidea
- (亜目)Stiripasterida
- (亜目)Eupantopodida
2010年代の現在でトックリウミグモ科とウスイロウミグモ科の種類は、形態に基づいた伝統的な分類体系ではそれぞれがイソウミグモ科とカニノテウミグモ科に含まれた[7]。しかし分子系統解析では、いずれも該当する科の種類とは遠縁の別系統だと示され[4][88]、後に区別された。カイヤドリウミグモなどを含んだカイヤドリウミグモ属(Nymphonella)に関しては、伝統的にユメムシ科もしくはイソウミグモ科に分類されたが[7]、分子系統解析ではトックリウミグモ科に含まれる[88][43]・カニノテウミグモ科とユメムシ科の姉妹群[87]・ミドリウミグモ科の姉妹群[87]など、様々な結果が出ている。
また、化石種のみ知られる科としてPalaeopycnogonididaeがあるが、現生科との類縁関係は不明確である[18]。
人間との係わり
編集通常は人間の日常生活とは関わりのない動物で、人間活動がウミグモ類に及ぼす影響も未解明である[46]。
なお、幼生が二枚貝に寄生する日本のカイヤドリウミグモ(Nymphonella tapetis)に関しては、2007年で東京湾の干潟において大量発生し、宿主とされるアサリの大量死に至って漁業被害を与えた[7][90][91]。ウミグモ類自体はなじみの薄い動物であるため、当時において本種は各メディアに「アサリに入っている変な虫」と報告された[7]。また、その性質により、漁業者たちから「海の吸血鬼」と呼ばれて忌み嫌われているとされている[45]。
脚注
編集注釈
編集- ^
- 左上:Austrodecus bamberi(スイクチウミグモ科)
- 右上:Colossendeis sp.(オオウミグモ科)
- 左中:Pycnogonum stearnsi(ヨロイウミグモ科)
- 右中:Ammothea hilgendorfi(イソウミグモ科)
- 左下:Endeis flaccida(ミドリウミグモ科)
- 右下:Nymphon signatum(ユメムシ科)
- ^ a b Decolopoda属、Pentacolossendeis属とPentanymphon属は5対、Dodecolopoda属は6対、Pentapycnon属は5-6対。
- ^ ヤマトトックリウミグモ(Ascorhynchus japonicus)・ハリエステス(Haliestes)などは4節、パレオイソプス(Palaeoisopus)は5節
- ^ a b 消化管の枝をもたない・肢節の数と構成が似る・プロトニンフォン幼生においては同形かつ同時に出揃うなどの類似性が指摘される。
- ^ Endeis laevis と Pycnogonum litorale の第2肢節生殖口は背側に開く。
- ^ Pycnogonum litorale の雌の生殖口は第4脚のみにある
- ^ a b ラディオドンタ類は文献によって前大脳のみ、もしくは前大脳と中大脳のみをもつとされる。
出典
編集- ^ a b c d e f Waloszek, Dieter; Dunlop, Jason A. (2002-05). “A Larval Sea Spider (Arthropoda: Pycnogonida) from the Upper Cambrian 'orsten' of Sweden, and the Phylogenetic Position of Pycnogonids” (英語). Palaeontology 45 (3): 421–446. doi:10.1111/1475-4983.00244. ISSN 0031-0239.
- ^ a b c d e Sabroux, Romain; Audo, Denis; Charbonnier, Sylvain; Corbari, Laure; Hassanin, Alexandre (2019-11-17). “150-million-year-old sea spiders (Pycnogonida: Pantopoda) of Solnhofen”. Journal of Systematic Palaeontology 17 (22): 1927–1938. doi:10.1080/14772019.2019.1571534. ISSN 1477-2019.
- ^ a b c d e 百科事典マイペディア,日本大百科全書(ニッポニカ). “ウミグモとは”. コトバンク. 2018年12月10日閲覧。
- ^ a b c d e f g h Arango, Claudia; Wheeler, W.C. (2007-06-01). “Phylogeny of the sea spiders (Arthropoda, Pycnogonida) based on direct optimization of six loci and morphology”. Cladistics 23: 255–293. doi:10.1111/j.1096-0031.2007.00143.x.
- ^ a b c d e f g h Hassanin, Alexandre; Prieur, Stéphanie le; Bonillo, Céline; Krapp, Franz; Corbari, Laure; Sabroux, Romain (2017-02-24). “Biodiversity and phylogeny of Ammotheidae (Arthropoda: Pycnogonida)” (英語). European Journal of Taxonomy 0 (286). doi:10.5852/ejt.2017.286. ISSN 2118-9773.
- ^ a b c Charbonnier, S.; Vannier, J.; Riou, B. (2007-10-22). “New sea spiders from the Jurassic La Voulte-sur-Rhône Lagerstätte”. Proceedings. Biological Sciences 274 (1625): 2555–2561. doi:10.1098/rspb.2007.0848. ISSN 0962-8452. PMC 2275891. PMID 17698484.
- ^ a b c d e f g h i j k 宮崎勝己、小林豊、鳥羽光晴、土屋仁「アサリに内部寄生し漁業被害を与えるカイヤドリウミグモの生物学」『タクサ:日本動物分類学会誌』第28号、2010年2月20日、45-54頁、doi:10.19004/taxa.28.0_45、ISSN 2189-7298。
- ^ a b c d e f g h i j k l m Dietz, Lars; Dömel, Jana S.; Leese, Florian; Lehmann, Tobias; Melzer, Roland R. (2018-03-15). “Feeding ecology in sea spiders (Arthropoda: Pycnogonida): what do we know?”. Frontiers in Zoology 15 (1): 7. doi:10.1186/s12983-018-0250-4. ISSN 1742-9994. PMC 5856303. PMID 29568315.
- ^ a b c d e f g h i j k l m n o p q r s t Bain, Bonnie; Govedich, Fredric (2004-12-01). “Courtship and mating behavior in the Pycnogonida (Chelicerata: Class Pycnogonida): A summary”. Invertebrate Reproduction & Development 46: 63–79. doi:10.1080/07924259.2004.9652607.
- ^ a b c d e f g h i j k l m n o Crooker, Allen (2008), Capinera, John L., ed. (英語), Sea Spiders (Pycnogonida), Springer Netherlands, pp. 3321–3335, doi:10.1007/978-1-4020-6359-6_4098, ISBN 978-1-4020-6359-6 2022年6月13日閲覧。
- ^ a b 中村光一郎「ウミグモ類」「皆脚亜門(ウミグモ亜門)」日高敏隆監修・奥谷喬司・武田正倫・今福道夫編『日本動物大百科 7 無脊椎動物』平凡社、1997年、122-123頁。
- ^ a b 小野展嗣「夢の虫」『クモ学 摩訶不思議な八本足の世界』東海大学出版会、2002年、115-118頁。
- ^ “Sea spiders provide insights into Antarctic evolution”. Department of the Environment and Energy, Australian Antarctic Division (22 July 2010). 27 December 2017閲覧。
- ^ a b c d e f g h i j k l m n o p q r s t u v w A., Dunlop, Jason; C., Lamsdell, James (2017-05). “Segmentation and tagmosis in Chelicerata” (英語). Arthropod Structure & Development 46 (3). ISSN 1467-8039.
- ^ a b c d e f g h i j k l m n o p q r s t u Bergstrom, J.; Stuermer, W.; Winter, G. (1980-07-20). “Palaeoisopus, Palaeopantopus and Palaeothea, pycnogonid arthropods from the Lower Devonian Hunsrück Slate, West Germany.” (英語). Palaeontologische Zeitschrift 54 (1-2): 7–54.
- ^ a b c d e f g h i Vilpoux, Kathia; Waloszek, Dieter (2003-12-01). “Larval development and morphogenesis of the sea spider Pycnogonum litorale (Ström, 1762) and the tagmosis of the body of Pantopoda” (英語). Arthropod Structure & Development 32 (4): 349–383. doi:10.1016/j.asd.2003.09.004. ISSN 1467-8039.
- ^ a b c d e f g h i j k l m n o Guyader, Hervé Le; Clabaut, Céline; Murienne, Jérôme; Jager, Muriel; Manuel, Michaël (2006-07-01). “Hox genes in sea spiders (Pycnogonida) and the homology of arthropod head segments” (英語). Development Genes and Evolution 216 (7-8): 481–491. doi:10.1007/s00427-006-0095-2. ISSN 1432-041X.
- ^ a b c d e f g Sabroux, Romain; Edgecombe, Gregory D.; Pisani, Davide; Garwood, Russell J. (2023-07). “New insights into the sea spider fauna (Arthropoda, Pycnogonida) of La Voulte‐sur‐Rhône, France (Jurassic, Callovian)” (英語). Papers in Palaeontology 9 (4). doi:10.1002/spp2.1515. ISSN 2056-2799.
- ^ a b c d e f g h i j k l 中村光一郎「真鶴海岸のウミグモ類」『横浜国立大学教育学部理科教育実習施設研究報告』第6巻、横浜国立大学教育学部附属理科教育実習施設、1990年3月25日、19–33頁。
- ^ a b c d e Siveter, David J.; Briggs, Derek E. G.; Sutton, Mark D.; Siveter, Derek J. (2004-10). “A Silurian sea spider” (英語). Nature 431 (7011): 978–980. doi:10.1038/nature02928. ISSN 1476-4687.
- ^ Fusco, Giuseppe; Minelli, Alessandro (2013), Minelli, Alessandro; Boxshall, Geoffrey, eds. (英語), Arthropod Segmentation and Tagmosis, Springer Berlin Heidelberg, pp. 197–221, doi:10.1007/978-3-642-36160-9_9, ISBN 978-3-642-36159-3 2022年4月16日閲覧。
- ^ a b c Thompson, Deborah P.; Young, Graham A.; Cuggy, Michael B.; Rudkin, David M. (2013/05). “An Ordovician Pycnogonid (Sea Spider) with Serially Subdivided ‘Head’ Region” (英語). Journal of Paleontology 87 (3): 395–405. doi:10.1666/12-057.1. ISSN 1937-2337.
- ^ “Marine Species Identification Portal : Macrobenthos of the North Sea - Pycnogonida : Glossary : cephalon”. species-identification.org. 2018年12月11日閲覧。
- ^ Miether, Sebastian T.; Dunlop, Jason A. (2016/07). “Lateral eye evolution in the arachnids”. Arachnology 17 (2): 103–119. doi:10.13156/arac.2006.17.2.103. ISSN 2050-9928.
- ^ a b Staples, David (2007-01-01). “A new species of Colossendeis (Pycnogonida: Colossendeidae) together with records from Australian and New Zealand waters”. Memoirs of Museum Victoria 64: 79–94. doi:10.24199/j.mmv.2007.64.8.
- ^ a b c d Sabroux, Romain; Garwood, Russell J.; Pisani, Davide; Donoghue, Philip C. J.; Edgecombe, Gregory D. (2024-10-14). “New insights into the Devonian sea spiders of the Hunsrück Slate (Arthropoda: Pycnogonida)” (英語). PeerJ 12: e17766. doi:10.7717/peerj.17766. ISSN 2167-8359.
- ^ a b Melzer, Roland R.; Leese, Florian; Hübner, Jeremy; Hofmann, Michaela; Dömel, Jana S.; Wagner, Philipp (2017-03-01). “Comparative study of bisected proboscides of Pycnogonida” (英語). Organisms Diversity & Evolution 17 (1): 121–135. doi:10.1007/s13127-016-0310-6. ISSN 1618-1077.
- ^ a b c d e Cano-Sánchez, Esperanza; López-González, Pablo J. (2016-12-15). “Basal articulation of the palps and ovigers in Antarctic Colossendeis (Pycnogonida; Colossendeidae)”. Helgoland Marine Research 70 (1): 22. doi:10.1186/s10152-016-0474-7. ISSN 1438-3888.
- ^ a b G. Rabay, Soraya; Matthews-Cascon, Helena; Bezerra, Luis (2017-04-24). “First record of Pentapycnon geayi Bouvier, 1911 (Pycnogonida: Pycnogonidae) in the state of Ceará, northeastern Brazil”. Check List 13: 2099. doi:10.15560/13.2.2099.
- ^ K, Miyazaki、T, Makioka「ウミグモ類(節足動物;ウミグモ類),Pentacolossendeis reticulata(オオウミグモ類)及びPentapycnon geayi(ウミグモ類)の余剰脚種における雌生殖孔に関するノート」『慶應義塾大学日吉紀要 自然科学』第16号、1994年、21–24頁、ISSN 0911-7237。
- ^ “Marine Species Identification Portal : Macrobenthos of the North Sea - Pycnogonida : Glossary : propodus”. species-identification.org. 2018年12月11日閲覧。
- ^ a b SHULTZ, JEFFREY W. (1989-09-01). “Morphology of locomotor appendages in Arachnida: evolutionary trends and phylogenetic implications”. Zoological Journal of the Linnean Society 97 (1): 1–56. doi:10.1111/j.1096-3642.1989.tb00552.x. ISSN 0024-4082.
- ^ a b c Poschmann, Markus; Dunlop, Jason A. (2006-09-01). “A New Sea Spider (arthropoda: Pycnogonida) with a Flagelliform ℡son from the Lower Devonian Hunsrück Slate, Germany” (英語). Palaeontology 49 (5): 983–989. doi:10.1111/j.1475-4983.2006.00583.x. ISSN 1475-4983.
- ^ “(PDF) Microscopic anatomy of Pycnogonida: II. Digestive system. III. Excretory system” (英語). ResearchGate. 2018年12月11日閲覧。
- ^ a b c d Moran, Amy L.; Arango, Claudia P.; Tobalske, Bret W.; Shishido, Caitlin; Lane, Steven J.; Woods, H. Arthur (2017-07-10). “Respiratory gut peristalsis by sea spiders” (English). Current Biology 27 (13): R638–R639. doi:10.1016/j.cub.2017.05.062. ISSN 0960-9822. PMID 28697358.
- ^ a b c d e Frankowski, Karina; Miyazaki, Katsumi; Brenneis, Georg (2022-03-31). “A microCT-based atlas of the central nervous system and midgut in sea spiders (Pycnogonida) sheds first light on evolutionary trends at the family level”. Frontiers in Zoology 19 (1): 14. doi:10.1186/s12983-022-00459-8. ISSN 1742-9994. PMC 8973786. PMID 35361245.
- ^ a b c d Brenneis, Georg; Ungerer, Petra; Scholtz, Gerhard (2008-11). “The chelifores of sea spiders (Arthropoda, Pycnogonida) are the appendages of the deutocerebral segment”. Evolution & Development 10 (6): 717–724. doi:10.1111/j.1525-142X.2008.00285.x. ISSN 1525-142X. PMID 19021742.
- ^ Brenneis, Georg; Scholtz, Gerhard (2014-04-15). “The ‘Ventral Organs’ of Pycnogonida (Arthropoda) Are Neurogenic Niches of Late Embryonic and Post-Embryonic Nervous System Development” (英語). PLOS ONE 9 (4): e95435. doi:10.1371/journal.pone.0095435. ISSN 1932-6203. PMC 3988247. PMID 24736377.
- ^ Woods, H. Arthur; Tobalske, Bret W.; Shishido, Caitlin M.; Moran, Amy L.; Lane, Steven J. (2018-04-15). “Cuticular gas exchange by Antarctic sea spiders” (英語). Journal of Experimental Biology 221 (8): jeb177568. doi:10.1242/jeb.177568. ISSN 1477-9145. PMID 29593081.
- ^ “【動画】ウミグモは「脚呼吸」、腸で酸素運ぶ”. natgeo.nikkeibp.co.jp. 2018年12月10日閲覧。
- ^ Munilla T, Soler-Membrives A. Check-list of the pycnogonids from Ant‑ arctic and sub-Antarctic waters: zoogeographic implications. Antarct Sci. 2009;21:99–111.
- ^ “The Biology of Pycnogonida” (英語). Advances in Marine Biology 24: 1–96. (1988-01-01). doi:10.1016/S0065-2881(08)60073-5. ISSN 0065-2881.
- ^ a b c d 張成年、丹羽健太郎、岡本俊治、村内嘉樹、平井玲、日比野学、涌井邦浩、冨山毅、小林豊、鳥羽光晴、狩野泰則「カイヤドリウミグモ Nymphonella tapetis 地域集団の遺伝的分化と分類学的位置」『日本水産学会誌』第78巻第5号、2012年、895–902頁、doi:10.2331/suisan.78.895、ISSN 1349-998X。
- ^ “Ascorhynchus cooki Child, 1987”. Museums Victoria Collections. 2018年12月11日閲覧。
- ^ a b 安田琢典 (2020年10月1日). “超レアなウミグモの捕食、担当者ものけぞる 鳥羽水族館”. 朝日新聞デジタル. 2020年10月9日閲覧。
- ^ a b c “南極のウミグモにくっつく生物、どれだけ重荷?”. 日経ナショナルジオグラフィック. 2018年12月10日閲覧。
- ^ Tomaschko, K.-H. (1994-07-01). “Ecdysteroids fromPycnogonum litorale (Arthropoda, Pantopoda) act as chemical defense againstCarcinus maenas (Crustacea, Decapoda)” (英語). Journal of Chemical Ecology 20 (7): 1445–1455. doi:10.1007/BF02059872. ISSN 1573-1561.
- ^ Brenneis, Georg; Frankowski, Karina; Maaß, Laura; Scholtz, Gerhard (2023-01-31). “The sea spider Pycnogonum litorale overturns the paradigm of the absence of axial regeneration in molting animals” (英語). Proceedings of the National Academy of Sciences 120 (5): e2217272120. doi:10.1073/pnas.2217272120. ISSN 0027-8424.
- ^ Lane, Steven J; Shishido, Caitlin M.; Moran, Amy L.; Tobalske, Bret W.; Lane, Steven J. (2018-08-01). “Costs of epibionts on Antarctic sea spiders” (英語). Marine Biology 165 (8): 137. doi:10.1007/s00227-018-3389-9. ISSN 1432-1793.
- ^ a b Burris, Zair P. (2011-02-01). “Costs of exclusive male parental care in the sea spider Achelia simplissima (Arthropoda: Pycnogonida)” (英語). Marine Biology 158 (2): 381–390. doi:10.1007/s00227-010-1566-6. ISSN 1432-1793.
- ^ Lehmann, Tobias; Gailer, Juan P.; Melzer, Roland R.; Schwabe, Enrico (2007-01-01). “A scanning-electron microscopic study of Dickdellia labioflecta (Dell, 1990) (Gastropoda, Littorinoidea) on Colossendeis megalonyx megalonyx Fry and Hedgpeth, 1969 (Pycnogonida, Colossendeidae): A test for ectoparasitism”. Polar Biology - POLAR BIOL 30: 243–248. doi:10.1007/s00300-006-0178-6.
- ^ Schiaparelli, Stefano; Oliverio, Marco; Taviani, Marco; Griffiths, Huw; Lörz, Anne-Nina; Albertelli, Giancarlo (2008). “Circumpolar distribution of the pycnogonid-ectoparasitic gastropod Dickdellia labioflecta (Dell, 1990) (Mollusca: Zerotulidae)” (英語). Antarct Sci 20 (5): 497–498. doi:10.1017/S0954102008001302. ISSN 0954-1020.
- ^ a b c Brenneis, Georg; Bogomolova, Ekaterina V.; Arango, Claudia P.; Krapp, Franz (2017-02-07). “From egg to “no-body”: an overview and revision of developmental pathways in the ancient arthropod lineage Pycnogonida”. Frontiers in Zoology 14 (1): 6. doi:10.1186/s12983-017-0192-2. ISSN 1742-9994. PMC 5297176. PMID 28191025.
- ^ 世界大百科事典内言及. “プロトニンフォン(ぷろとにんふぉん)とは”. コトバンク. 2018年12月11日閲覧。
- ^ Alexeeva, Nina; Tamberg, Yuta; Shunatova, Natalia (2018-05-01). “Postembryonic development of pycnogonids: A deeper look inside” (英語). Arthropod Structure & Development 47 (3): 299–317. doi:10.1016/j.asd.2018.03.002. ISSN 1467-8039.
- ^ a b c d Kühl, Gabriele; Poschmann, Markus; Rust, Jes (2013-05-01). “A ten-legged sea spider (Arthropoda: Pycnogonida) from the Lower Devonian Hunsruck Slate (Germany)”. Geological Magazine online. doi:10.1017/S0016756812001033.
- ^ Bamber, Roger N. (2007-12-21). “A holistic re-interpretation of the phylogeny of the Pycnogonida Latreille, 1810 (Arthropoda)*” (英語). Zootaxa 1668 (1): 295–312. doi:10.11646/zootaxa.1668.1.15. ISSN 1175-5334.
- ^ Edgecombe, Gregory D. (2010-3). “Arthropod phylogeny: an overview from the perspectives of morphology, molecular data and the fossil record”. Arthropod Structure & Development 39 (2-3): 74–87. doi:10.1016/j.asd.2009.10.002. ISSN 1873-5495. PMID 19854297.
- ^ Siveter, Derek J.; Sabroux, Romain; Briggs, Derek E. G.; Siveter, David J.; Sutton, Mark D. (2023-09). “Newly discovered morphology of the Silurian sea spider Haliestes and its implications” (英語). Papers in Palaeontology 9 (5). doi:10.1002/spp2.1528. ISSN 2056-2799.
- ^ Poschmann, Markus; Dunlop, Jason A. (2006-09). “A new sea spider (Arthropoda: Pycnogonida) with a flagelliform telson from the Lower Devonian Hunsrück Slate, Germany” (英語). Palaeontology 49 (5): 983–989. doi:10.1111/j.1475-4983.2006.00583.x.
- ^ a b c Maxmen, Amy; Browne, William E.; Martindale, Mark Q.; Giribet, Gonzalo (2005). “Neuroanatomy of sea spiders implies an appendicular origin of the protocerebral segment”. Nature 437 (7062): 1144–8. Bibcode: 2005Natur.437.1144M. doi:10.1038/nature03984. PMID 16237442.
- ^ a b Jager, Muriel; Murienne, Jérôme; Clabaut, Céline; Deutsch, Jean; Guyader, Hervé Le; Manuel, Michaël (2006). “Homology of arthropod anterior appendages revealed by Hox gene expression in a sea spider”. Nature 441 (7092): 506–8. Bibcode: 2006Natur.441..506J. doi:10.1038/nature04591. PMID 16724066.
- ^ a b Dunlop, J. A.; Arango, C. P. (2005-02-01). “Pycnogonid affinities: a review” (英語). Journal of Zoological Systematics and Evolutionary Research 43 (1): 8–21. doi:10.1111/j.1439-0469.2004.00284.x. ISSN 1439-0469.
- ^ Vlášková, M.; Hypša, V.; Zrzavý, J. (1998) (英語). Arthropod Relationships. Springer, Dordrecht. pp. 97–107. doi:10.1007/978-94-011-4904-4_9. ISBN 9789401060578
- ^ Wheeler, Ward C.; Edgecombe, Gregory D.; Giribet, Gonzalo (2001-09). “Arthropod phylogeny based on eight molecular loci and morphology” (英語). Nature 413 (6852): 157–161. doi:10.1038/35093097. ISSN 1476-4687.
- ^ Legg, David (2013-06) (英語). The impact of fossils on arthropod phylogeny. doi:10.25560/24168.
- ^ Edgecombe, Gregory D.; Wilson, George D. F.; Colgan, Donald J.; Gray, Michael R.; Cassis, Gerasimos (2000-06-01). “Arthropod Cladistics: Combined Analysis of Histone H3 and U2 snRNA Sequences and Morphology” (英語). Cladistics 16 (2): 155–203. doi:10.1111/j.1096-0031.2000.tb00352.x. ISSN 1096-0031.
- ^ Roeding, Falko; Borner, Janus; Kube, Michael; Klages, Sven; Reinhardt, Richard; Burmester, Thorsten (2009-12-01). “A 454 sequencing approach for large scale phylogenomic analysis of the common emperor scorpion (Pandinus imperator)” (英語). Molecular Phylogenetics and Evolution 53 (3): 826–834. doi:10.1016/j.ympev.2009.08.014. ISSN 1055-7903.
- ^ a b Regier, Jerome C.; Shultz, Jeffrey W.; Zwick, Andreas; Hussey, April; Ball, Bernard; Wetzer, Regina; Martin, Joel W.; Cunningham, Clifford W. (2010-02). “Arthropod relationships revealed by phylogenomic analysis of nuclear protein-coding sequences” (英語). Nature 463 (7284): 1079–1083. doi:10.1038/nature08742. ISSN 0028-0836.
- ^ a b Rehm, Peter; Meusemann, Karen; Borner, Janus; Misof, Bernhard; Burmester, Thorsten (2014-08-01). “Phylogenetic position of Myriapoda revealed by 454 transcriptome sequencing” (英語). Molecular Phylogenetics and Evolution 77: 25–33. doi:10.1016/j.ympev.2014.04.007. ISSN 1055-7903.
- ^ a b Sharma, Prashant P.; Kaluziak, Stefan T.; Pérez-Porro, Alicia R.; González, Vanessa L.; Hormiga, Gustavo; Wheeler, Ward C.; Giribet, Gonzalo (2014-11-01). “Phylogenomic Interrogation of Arachnida Reveals Systemic Conflicts in Phylogenetic Signal” (英語). Molecular Biology and Evolution 31 (11): 2963–2984. doi:10.1093/molbev/msu235. ISSN 0737-4038.
- ^ “The Phylogeny and Evolutionary History of Arthropods” (英語). Current Biology 29 (12): R592–R602. (2019-06-17). doi:10.1016/j.cub.2019.04.057. ISSN 0960-9822.
- ^ Edgecombe, Gregory D. (2020-07-31). “Arthropod Origins: Integrating Paleontological and Molecular Evidence”. Annual Review of Ecology, Evolution, and Systematics. doi:10.1146/annurev-ecolsys-011720-124437. ISSN 1543-592X.
- ^ Giribet, G (2002-02). “Phylogeny and Systematic Position of Opiliones: A Combined Analysis of Chelicerate Relationships Using Morphological and Molecular Data”. Cladistics 18 (1): 5–70. doi:10.1006/clad.2001.0185. ISSN 0748-3007.
- ^ Hassanin, Alexandre; Léger, Nelly; Deutsch, Jean (2005-05-01). “Evidence for Multiple Reversals of Asymmetric Mutational Constraints during the Evolution of the Mitochondrial Genome of Metazoa, and Consequences for Phylogenetic Inferences”. Systematic biology 54: 277–98. doi:10.1080/10635150590947843.
- ^ Masta, Susan E.; McCall, Andrew; Longhorn, Stuart J. (2010-10-01). “Rare genomic changes and mitochondrial sequences provide independent support for congruent relationships among the sea spiders (Arthropoda, Pycnogonida)” (英語). Molecular Phylogenetics and Evolution 57 (1): 59–70. doi:10.1016/j.ympev.2010.06.020. ISSN 1055-7903.
- ^ Hassanin, Alexandre (2006-01-01). “Phylogeny of Arthropoda inferred from mitochondrial sequences: Strategies for limiting the misleading effects of multiple changes in pattern and rates of substitution” (英語). Molecular Phylogenetics and Evolution 38 (1): 100–116. doi:10.1016/j.ympev.2005.09.012. ISSN 1055-7903.
- ^ Aria, Cédric; Caron, Jean-Bernard (2019-09-26). “A middle Cambrian arthropod with chelicerae and proto-book gills” (英語). Nature 573 (7775): 586–589. doi:10.1038/s41586-019-1525-4. ISSN 0028-0836.
- ^ “PycnoBase - Pycnogonida”. www.marinespecies.org. 2018年12月10日閲覧。
- ^ a b 花井哲郎・山口寿之・冨田幸光・尾崎公彦・植村和彦「生物分類表」日本古生物学会編『古生物学事典』朝倉書店、1991年、348-363頁。
- ^ a b 文部省・日本動物学会編「動物分類名」『学術用語集 動物学編(増訂版)』丸善、1988年、1060-1100頁。
- ^ a b c d 小野展嗣「鋏角亜門分類表」石川良輔編『節足動物の多様性と系統』〈バイオディバーシティ・シリーズ〉6、岩槻邦男・馬渡峻輔監修、裳華房、2008年、410-420頁。
- ^ a b ジョヴァンニ・ピンナ「海蜘蛛(ウミグモ)上綱(Pycnogonida)」小畠郁生監訳・二上政夫訳『図解 世界の化石大百科』河出書房新社、2000年、102-103頁。
- ^ “PycnoBase - Palaeoisopodus problematicus Boili, 1928 †”. www.marinespecies.org. 2018年12月10日閲覧。
- ^ “PycnoBase - Nectopantopoda †”. www.marinespecies.org. 2018年12月10日閲覧。
- ^ “Haliestes dasos |” (英語). 2018年12月10日閲覧。
- ^ a b c d e Ballesteros, Jesús A.; Setton, Emily V. W.; López, Carlos E. Santibáñez; Arango, Claudia P.; Brenneis, Georg; Brix, Saskia; Cano-Sánchez, Esperanza; Dandouch, Merai et al. (2020-02-02). “Phylogenomic resolution of sea spider diversification through integration of multiple data classes” (英語). bioRxiv: 2020.01.31.929612. doi:10.1101/2020.01.31.929612.
- ^ a b c d e Hassanin, Alexandre; Couloux, Arnaud; Cruaud, Corinne; Arabi, Juliette. “Studying sources of incongruence in arthropod molecular phylogenies: Sea spiders (Pycnogonida) as a case study” (英語). Comptes Rendus Biologies 333 (5): 438–453. ISSN 1631-0691.
- ^ “PycnoBase - Pycnogonida”. www.marinespecies.org. 2018年12月10日閲覧。
- ^ “カイヤドリウミグモ”. 2018年12月16日閲覧。
- ^ “カイヤドリウミグモ(1)”. marine1.bio.sci.toho-u.ac.jp. 2018年12月16日閲覧。