ディバステリウム
ダイバステリウム[2](Dibasterium)は、約4億2,500万年前のシルル紀に生息した化石節足動物の一属[1]。たくさんの肢を生えたカブトガニに似た鋏角類であり[3]、イギリスのヘレフォードシャー保存堆積地(Herefordshire Konservat-Lagerstätte)で見つかった Dibasterium durgae という1種のみによって知られる[4][5]。
| ダイバステリウム | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
 ダイバステリウムの復元図
| ||||||||||||||||||
| 地質時代 | ||||||||||||||||||
| 古生代シルル紀(約4億2,500万年前)[1] | ||||||||||||||||||
| 分類 | ||||||||||||||||||
| ||||||||||||||||||
| 学名 | ||||||||||||||||||
| Dibasterium Briggs et al., 2012[1] | ||||||||||||||||||
| タイプ種 | ||||||||||||||||||
| Dibasterium durgae Briggs et al., 2012[1] |
二叉型の脚は歩脚状の外肢をもつことによって、一見では通常の鋏角類より多くの手足が生えたように見える。学名「Dibasterium」はその特徴に因んで、ラテン語の「dibamos」(二足)と「mysterium」(神秘)の合成語で、模式種(タイプ種)の種小名「durgae」はたくさんの腕をもつヒンドゥー教の女神ドゥルガー(Durga)に由来する[1]。オックスフォード大学自然史博物館(Oxford University Museum of Natural History)に所蔵される唯一の化石標本(OUMNH C.29640)のみが記載される[1][4]。
形態
編集-
 ダイバステリウムの全身復元図
ダイバステリウムの全身復元図 -
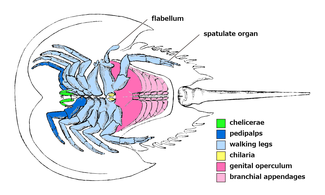
 ダイバステリウムの付属肢(1:鋏角、2-6:脚(内肢)、7:唇様肢らしき付属肢、8-13:蓋板、Ex:脚の外肢)
ダイバステリウムの付属肢(1:鋏角、2-6:脚(内肢)、7:唇様肢らしき付属肢、8-13:蓋板、Ex:脚の外肢)

体長2.3cmで横幅1cm程度の、縦に長いカブトガニのような姿をした小型節足動物である[1]。体は丸い背甲に覆われる前体と分節した後体に分かれ、後体はさらに幅広い前腹部と細長い後腹部に区別される[1]。付属肢(関節肢)は体の腹面に隠れる[1]。
以下の前体と後体の区分は Dunlop & Lamsdell 2017 および鋏角類の通常の体制(第1-6体節/付属肢対は前体で、後体はそれ以降)に基づく[3]。
前体
編集前体(prosoma)は先節と第1-6体節の癒合でできた合体節とされ[3]、ドーム型の丸い背甲(prosomal dorsal shield)に覆われる。背甲の表面は没個性で、眼や隆起線などの構造は見当たらない[1]。前後で1対の鋏角と5対の脚という計6対の付属肢があり[3]、脚は外肢を除いてその全ての付け根は腹面中央(口の周辺)に集約する[1]。前4対の脚の外肢は歩脚状に発達するため、まるで9対の脚が生えたように見える[1]。
鋏角(chelicera)の先端は多くの鋏角類と同じくはさみ型だが、柄部は異様に細長く湾曲しており、おそらく複数の肢節に分かれたと考えられる[1]。
5対の脚のうち前の4対は外肢(exopod)と内肢(endopod)をもつ二叉型で、最終の1対は内肢のみをもつ単枝型である[1][3]。それぞれの脚の基部に顎基(gnathobase)がある[1]。内肢は基部を除いて7つの肢節からなり、そのうち前の4対は先端2節がはさみをなし、最終の1対は先端数節にたくさんの棘が生える[1]。4対の外肢は内肢よりやや短い歩脚状で、5つの肢節に分かれ、そのうち先端数節のそれぞれの先端に長い刺毛が生える(第3肢節1本、第4肢節5本、第5肢節3本)[1]。これらの外肢の接続部は体壁を介して、完全に内肢から分離していたと考えている[1][3]。
後体
編集後体(opisthosoma)は明瞭に分節した11節(第7-17体節)からなり、前後でさらに8節の前腹部(preabdomen)と3節の後腹部(postabdomen)に区別される[1]。前腹部の各体節は盛り上がった背板(tergite)に覆われ、そのうち第1節の背板は退化的で、前半部が直前の背甲に覆われる[1]。残り7節の背板は発達して1対の溝が背面を走り、左右に出っ張り(pleurae)があって後方ほど尖る[1]。後腹部の3節は前腹部より幅が狭い環状で、左右は小さな棘が生える程度で前腹部のような出っ張りはない[1]。後端の尾節(telson)は剣状で三角形の断面をしており、先端がわずかに二股に分かれる[1]。尾節と最終体節の腹面の接続部に肛門がある[1]。
後体はその前7節(第7-13体節)に由来する7対の付属肢がある[1][3]。最初の1対は脚の直後に配置される小さな付属肢であり、刺毛が生えた葉状で、カブトガニ類の唇様肢(chilarium)を思わせる[1][5]。残り6対の付属肢はカブトガニ類にも見られるような鰭状の蓋板(operculum)であり、左右の先端はやや重なっている[1]。しかしカブトガニ類などとは異なり、ダイバステリウムはほぼ同形の全ての蓋板に書鰓(book gill)をもつ[1][5](カブトガニ類とウミサソリ類の最初の蓋板は生殖口蓋 (genital operculum) としてやや特化し、書鰓を欠く[5])。
系統関係
編集
| |||||||||||||||||||||||||||||||||||||||
| 化石群(†)を中心とし、簡略化した鋏角類の内部系統におけるダイバステリウムの位置[6][7][8][9][10][11]。系統位置に諸説があるものは複数分岐としてまとめられる。 |
)_(20928914062).jpg)




.png)

ダイバステリウム属(Dibasterium)は Dibasterium durgae という1種のみによって知られる[5]。原記載では、ダイバステリウムはハラフシカブトガニ類と見なされ[1]、当時においてハラフシカブトガニ類は系統的にカブトガニ類に含まれた(=広義のカブトガニ類)[12]。しかしその原記載が公表された直後、ハラフシカブトガニ類の系統関係に対する再検討がなされ、そのほとんどが他のカブトガニ類とは直接的に類縁でないものと見なされるようになり(詳細はハラフシカブトガニ類#分類と進化を参照)、系統的他のカブトガニ類(=狭義のカブトガニ類)から区別され、ダイバステリウムとカブトガニ類の直接的な類縁関係もそれによって否定的に評価された[12]。それ以降では便宜的にハラフシカブトガニ類扱いされる場合もある[6][7][11]が、後述の形質に因んで、ダイバステリウムはオファコルスと共に、基盤的な真鋏角類であることはほとんどの系統解析に支持される[6][7][8][9][10][11]。
-
-
 ダイバステリウムのように歩脚状の4対の外肢(Ex)をもつオファコルス
ダイバステリウムのように歩脚状の4対の外肢(Ex)をもつオファコルス -
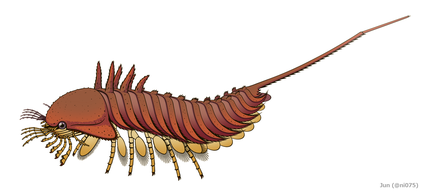



ダイバステリウムとオファコルスの多くの特徴(発達した背甲・腹面中央に集約する脚・はさみのある第1-4脚・能動的な6対の蓋板など)はカブトガニ類に共通し、特にダイバステリウムの体節区分と付属肢構造(脚の顎基・はさみのない第5脚・唇様肢らしき付属肢をもつなど)はオファコルス以上にカブトガニ類的である[1][3]。一方で、この2属の第1-4脚にある歩脚状の外肢は、カブトガニ類・ウミサソリ類・クモガタ類など他の真鋏角類に見当たらず、むしろ基盤的な鋏角類とされるカンブリア紀のハベリア類(ハベリア、サンクタカリスなど)を思わせる特徴である[1][13][9][10]。ダイバステリウムが6対の書鰓をもつことも、それが5対以下のみをもつ前述の真鋏角類とは異なる[14][3]。
これらの性質に基づいて、ダイバステリウムはオファコルスと共に多くの系統解析において基盤的な真鋏角類と見なされる[6][7][8][9][10][11]。これにより、基盤的な真鋏角類はかつて第1-4脚に歩脚状の外肢と後体に6対の書鰓を有し、それより派生的な真鋏角類の系統(Prosomapoda)に至って外肢が退化消失し[1][12][13][9][10]、書鰓/書肺の数も減少していたと考えられる[14][15][12]。ダイバステリウムの第1-4脚の外肢が体壁を介して内肢から分離した性質は、外肢と内肢が基部に繋がった原始的な二叉型付属肢と、外肢を失った単枝型付属肢の中間形態を表した特徴であると考えられる[1][4]。
脚注
編集- ^ a b c d e f g h i j k l m n o p q r s t u v w x y z aa ab ac ad ae af ag ah Briggs, Derek E. G.; Siveter, Derek J.; Siveter, David J.; Sutton, Mark D.; Garwood, Russell J.; Legg, David (2012-09-25). “Silurian horseshoe crab illuminates the evolution of arthropod limbs” (英語). Proceedings of the National Academy of Sciences 109 (39): 15702–15705. doi:10.1073/pnas.1205875109. ISSN 0027-8424. PMC 3465403. PMID 22967511.
- ^ 小林快次 (2014) (Japanese). 大昔の生きもの. 東京: ポプラ社. ISBN 978-4-591-14071-0. OCLC 883613863
- ^ a b c d e f g h i Dunlop, Jason A.; Lamsdell, James C. (2017). “Segmentation and tagmosis in Chelicerata” (英語). Arthropod Structure & Development 46 (3): 395. ISSN 1467-8039.
- ^ a b c Siveter, Derek J.; Briggs, Derek E. G.; Siveter, David J.; Sutton, Mark D. (2019). “The Herefordshire Lagerstätte: fleshing out Silurian marine life” (英語). Journal of the Geological Society 177 (1): 1–13. doi:10.1144/jgs2019-110. ISSN 0016-7649.
- ^ a b c d e Dunlop, J. A., Penney, D. & Jekel, D. 2020. A summary list of fossil spiders and their relatives. In World Spider Catalog. Natural History Museum Bern, online at http://wsc.nmbe.ch, version 20.5
- ^ a b c d Selden, Paul A.; Lamsdell, James C.; Qi, Liu (2015). “An unusual euchelicerate linking horseshoe crabs and eurypterids, from the Lower Devonian (Lochkovian) of Yunnan, China” (英語). Zoologica Scripta 44 (6): n/a–n/a. ISSN 0300-3256.
- ^ a b c d Lamsdell, James C.; Briggs, Derek E. G.; Liu, Huaibao P.; Witzke, Brian J.; McKay, Robert M. (2015-10). “A new Ordovician arthropod from the Winneshiek Lagerstätte of Iowa (USA) reveals the ground plan of eurypterids and chasmataspidids” (英語). The Science of Nature 102 (9-10): 63. doi:10.1007/s00114-015-1312-5. ISSN 0028-1042.
- ^ a b c Lamsdell, James C. (2016). “Horseshoe crab phylogeny and independent colonizations of fresh water: ecological invasion as a driver for morphological innovation” (英語). Palaeontology 59 (2): 181–194. doi:10.1111/pala.12220. ISSN 1475-4983.
- ^ a b c d e Aria, Cédric; Caron, Jean-Bernard (2017-12-21). “Mandibulate convergence in an armoured Cambrian stem chelicerate”. BMC Evolutionary Biology 17 (1): 261. doi:10.1186/s12862-017-1088-7. ISSN 1471-2148. PMC 5738823. PMID 29262772.
- ^ a b c d e Aria, Cédric; Caron, Jean-Bernard (2019-09). “A middle Cambrian arthropod with chelicerae and proto-book gills” (英語). Nature 573 (7775): 586–589. doi:10.1038/s41586-019-1525-4. ISSN 1476-4687.
- ^ a b c d Bicknell, Russell D.C.; Lustri, Lorenzo; Brougham, Tom (2019-12). “Revision of “Bellinurus” carteri (Chelicerata: Xiphosura) from the Late Devonian of Pennsylvania, USA” (英語). Comptes Rendus Palevol 18 (8): 967–976. doi:10.1016/j.crpv.2019.08.002.
- ^ a b c d Lamsdell, James C. (2013-01). “Revised systematics of Palaeozoic ‘horseshoe crabs’ and the myth of monophyletic Xiphosura: Re-evaluating the Monophyly of Xiphosura” (英語). Zoological Journal of the Linnean Society 167 (1): 1–27. doi:10.1111/j.1096-3642.2012.00874.x.
- ^ a b Legg, David A. (2014-12). “Sanctacaris uncata: the oldest chelicerate (Arthropoda)” (英語). Naturwissenschaften 101 (12): 1065–1073. doi:10.1007/s00114-014-1245-4. ISSN 0028-1042.
- ^ a b Dunlop, Jason (1998) (英語). The origins of tetrapulmonate book lungs and their significance for chelicerate phylogeny.
- ^ Scholtz, Gerhard; Kamenz, Carsten (2006). “The book lungs of Scorpiones and Tetrapulmonata (Chelicerata, Arachnida): evidence for homology and a single terrestrialisation event of a common arachnid ancestor”. Zoology (Jena, Germany) 109 (1): 2–13. doi:10.1016/j.zool.2005.06.003. ISSN 0944-2006. PMID 16386884.