リンゴ酸デヒドロゲナーゼ
リンゴ酸デヒドロゲナーゼまたはリンゴ酸脱水素酵素(malate dehydrogenase, MDH)は、リンゴ酸とオキサロ酢酸との相互変換を触媒する酸化還元酵素である。
-
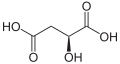 (S)-リンゴ酸・L-リンゴ酸
(S)-リンゴ酸・L-リンゴ酸 -
 オキサロ酢酸
オキサロ酢酸


用いる電子受容体によって以下の表のように分類されているほか、さらに脱炭酸してピルビン酸にするリンゴ酸酵素に対しても「リンゴ酸デヒドロゲナーゼ」と呼ぶことがある。
| IUBMB名称 | EC番号 | 電子受容体 |
|---|---|---|
| リンゴ酸デヒドロゲナーゼ | 1.1.1.37 | NAD+ |
| リンゴ酸デヒドロゲナーゼ (NADP+) | 1.1.1.82 | NADP+ |
| リンゴ酸デヒドロゲナーゼ [NAD(P)+] | 1.1.1.299 | NAD(P)+ |
| リンゴ酸デヒドロゲナーゼ (キノン) | 1.1.5.4 | キノン |
本項ではNAD+依存型の酵素(EC 1.1.1.37)について記述する。

リンゴ酸デヒドロゲナーゼまたはリンゴ酸脱水素酵素(malate dehydrogenase, MDH)は、リンゴ酸をオキサロ酢酸へと酸化する(またはその逆の)化学反応を触媒する酸化還元酵素である。クエン酸回路を構成する酵素の1つであり、また真核生物においてはリンゴ酸-アスパラギン酸シャトルに関与している。
- (S)-リンゴ酸 + NAD+ オキサロ酢酸 + NADH + H+

機能
編集- クエン酸回路
- しばしばクエン酸回路の出発物質の1つと目されるオキサロ酢酸を、リンゴ酸を酸化することで再生する働きを担っている。このときNAD+が還元されて、還元等量であるNADHを生じる。
- リンゴ酸-アスパラギン酸シャトル
- 細胞質における異化によって生じたNADHは、NAD+へと再酸化する必要がある。ミトコンドリアの電子伝達系はNADHの再酸化と共役させることで効率的にATP合成を行えるが、NADHは直接ミトコンドリア内膜を透過することができない。そこでリンゴ酸デヒドロゲナーゼとアスパラギン酸アミノ基転移酵素を組み合わせ、間接的にNADHをミトコンドリアへと輸送している。
- 糖新生
- とくに動物細胞においては糖新生にも重要である。糖以外の炭素骨格はミトコンドリア内でオキサロ酢酸かさらにホスホエノールピルビン酸(PEP)に変換され、これが細胞質での糖新生の出発物質となる。PEPはミトコンドリア内膜を透過できるが、しかしオキサロ酢酸の場合は動物細胞のミトコンドリア内膜を透過できない。そこでいったんリンゴ酸かアスパラギン酸に変えてミトコンドリアから運び出す必要がある。細胞質での糖新生には還元等量としてNADHが必要なので、たいていの場合はリンゴ酸を経由することでNADHも同時に細胞質へ輸送することになる。
アイソザイム
編集ヒトや他の多くの哺乳動物は以下の2種のリンゴ酸デヒドロゲナーゼを持つ。MDH1は細胞質に存在し、MDH2はミトコンドリアに存在する。
|
| ||||||||||||||||||||||||||||||||||||||||||||
構造と進化
編集リンゴ酸デヒドロゲナーゼ(MDH)の翻訳産物は分子量30~40kDa程度であるが、これがホモ2量体またはホモ4量体の四次構造をとる。MDHの祖先はホモ4量体であったと考えられ、構造上よく似た乳酸脱水素酵素もホモ4量体である。古細菌はホモ4量体のMDHを持っており、真正細菌でもグラム陽性菌やαプロテオバクテリアなどが該当する。これに対しほとんどの真核生物はホモ2量体のMDHを持っている。ホモ2量体のMDHは大きく分けて2種類あって、哺乳類では細胞質型のMDH1とミトコンドリア型のMDH2という様に区別されている。MDH1は、真核生物の細胞質や葉緑体で機能しているほか、真正細菌の一部(抗酸菌や放線菌など)が持っている。MDH2は真核生物のミトコンドリアで機能している以外に、キネトプラスト類のグリコソームや、真正細菌のγプロテオバクテリアなどが持っている。[1]
参考文献
編集- ^ Madern (2002). “Molecular evolution within the L-malate and L-lactate dehydrogenase super-family”. J. Mol. Evol. 54 (6): 825-840. doi:10.1007/s00239-001-0088-8.
- Banaszak, LJ, Bradshaw RA (1975). “Malate dehydrogenase”. In Boyer PD. The Enzymes. 11 (3rd ed.). New York: Academic Press. pp. 369–396
- Guha A, Englard S, Listowsky I (February 1968). “Beef heart malic dehydrogenases. VII. Reactivity of sulfhydryl groups and conformation of the supernatant enzyme”. J. Biol. Chem. 243 (3): 609–15. PMID 5637713.
- McReynolds MS, Kitto GB (February 1970). “Purification and properties of Drosophila malate dehydrogenases”. Biochim. Biophys. Acta 198 (2): 165–75. PMID 4313528.
- Wolfe RG, Nielands JB (July 1956). “Some molecular and kinetic properties of heart malic dehydrogenase”. J. Biol. Chem. 221 (1): 61–9. PMID 13345798.
外部リンク
編集- 糖新生の経路
- Malate dehydrogenase - MeSH・アメリカ国立医学図書館・生命科学用語シソーラス