行動遺伝学
行動遺伝学(こうどういでんがく、英: Behavioural genetics)とは、行動における個人差の本質と起源を調査するために遺伝学的手法を用いる科学的研究分野である。 「行動遺伝学」という名称は遺伝的影響に焦点を当てていることを示唆しているが、この分野では、遺伝的要因と環境的要因が個人差にどの程度影響を与えているのか、また、遺伝子と環境の交絡を取り除くことができる研究デザインの開発を広く研究している。行動遺伝学は、19世紀後半にフランシス・ゴルトンによって科学的分野として設立されたが、第二次世界大戦の前後に優生学運動と関連付けられたことで信用を失った。20世紀後半になると、この分野は、人間の行動と精神疾患の遺伝に関する研究(典型的には双生児と家族の研究を使用)、および選択的交配と交配による遺伝的に有益なモデル生物の研究によって、再び注目されるようになった。20世紀後半から21世紀初頭にかけて、分子遺伝学の技術的進歩により、ゲノムを直接測定し、修飾することが可能になった。これにより、モデル生物の研究(例えばノックアウトマウス)や人間の研究(例えばゲノムワイド関連研究)で大きな進歩があり、新しい科学的発見につながった。
行動遺伝学研究の知見は、行動に対する遺伝的および環境的影響の役割に関する現代の理解に広く影響を与えている。これには、研究されたほとんどすべての行動が、かなりの程度の遺伝的影響を受けているという証拠や、その影響は個人が成人になるにつれて増加する傾向があるという証拠が含まれる。さらに、研究された人間の行動のほとんどは、非常に多数の遺伝子の影響を受けており、これらの遺伝子の個々の影響は非常に小さい。環境的影響も強い役割を果たしているが、それらは家族を互いに似ているようにするのではなく、互いに異なるようにする傾向がある。
歴史
編集
選択的育種と動物の家畜化は、行動の個人差が自然な原因によるものかもしれないという考えを人類が持っていた最も古い証拠かもしれない[1]。プラトンとアリストテレスはそれぞれ、行動特性の遺伝の基礎とメカニズムについて推測した[2]。例えば、プラトンは、『国家』の中で、理想的な社会の追求において、ある特性の発達を奨励し、他の特性を阻止するために、市民の間で選択的育種を奨励すべきであると論じたが、今日では優生学と呼ばれるかもしれない[2][3]。行動遺伝学の概念は、イギリス・ルネサンスの時代にも存在しており、ウィリアム・シェイクスピアがテンペストの中で、「生まれか育ちか」という言葉を初めて作ったのかもしれない。そこでは、第4幕第1場で、キャリバンについて、「悪魔だ、生まれついての悪魔だ、その本性に教育は決して染みつかない」と書いている[3][4]。
現代の行動遺伝学は、19世紀の知識人でチャールズ・ダーウィンのいとこであるサー・フランシス・ゴルトンから始まった[3]。ゴルトンは博学者であり、人間の能力と精神的特性の遺伝を含む多くの分野を研究した。ゴルトンの研究の1つは、イギリスの上流階級における社会的・知的達成度に関する大規模な系図研究であった。1869年、ダーウィンの種の起源から10年後、ゴルトンはHereditary Genius誌でその結果を発表した[5]。この研究で、ゴルトンは、「著名人」の割合は著名人の近親者の間で最も高く、著名人との関係の度合いが減少するにつれて減少することを発見した。ゴルトンは、著名さに対する環境的影響の役割を否定できなかったが(このことは彼自身が認めていた)、この研究は、行動特性に対する遺伝と環境の相対的役割についての重要な議論のきっかけとなった。ゴルトンは、自身の研究を通じて、「多変量解析を導入し、現代のベイズ統計学への道を開いた」のであり、これは科学全体で使用されている—「統計学の啓蒙」と呼ばれるものを開始したのである[6]。

ゴルトンによって設立された行動遺伝学の分野は、最終的に、ゴルトンの知的貢献のもう1つである、20世紀社会における優生学運動の設立によって弱体化された[3]。優生学の主な考え方は、選択的育種と行動の遺伝に関する知識を組み合わせて、人類を改良することであった[3]。優生学運動は、その後、ナチス・ドイツにおける科学的な腐敗と大量虐殺行為によって信用を失った。行動遺伝学は、優生学との関連性から信用を失った[3]。この分野は、カルビン・S・ホールの1951年の行動遺伝学に関する本の章など、行動遺伝学に関する初期のテキストの出版を通じて、再び独立した科学分野としての地位を得た。この章で彼は、1960年代から1970年代にかけてある程度の人気を博した「精神遺伝学」という用語を導入した[7][8][9]。しかし、最終的には「行動遺伝学」という用語に取って代わられた。
行動遺伝学が明確な分野として始まったのは、1960年にジョン・L・フラーとウィリアム・ロバート(ボブ)・トンプソンによって Behavior Genetics という本が出版されたことによって示された[1][10]。現在では、動物や人間の行動の多く、あるいはほとんどが、重要な遺伝的影響を受けていることが広く認められているが、特定の形質に対する遺伝的影響の程度は大きく異なる可能性がある[11][12]。それから10年後の1970年2月、Behavior Genetics誌の第1号が発行され、1972年には行動遺伝学会が設立され、テオドシウス・ドブジャンスキーが初代会長に選出された。その後、この分野は成長し、多様化し、多くの科学分野に影響を与えている[3][13]。
方法
編集行動遺伝学の主な目的は、行動における個人差の本質と起源を調査することである[3]。行動遺伝学研究では、さまざまな方法論的アプローチが用いられているが[14]、そのうちのいくつかを以下に概説する。
動物研究
編集この節の加筆が望まれています。 |
動物行動遺伝学の研究者は、環境要因を慎重に制御し、遺伝的変異を実験的に操作することができるため、ヒトの行動遺伝学の研究では得られない程度の因果推論が可能である[15]。動物研究では、選択実験がよく用いられる。例えば、実験室のハツカネズミは、オープンフィールド行動[16]、体温調節の巣作り本能[17]、自発的な回し車運動行動[18]などについて選択交配されている。これらのデザインにおける一連の方法については、それぞれのページで説明されている。 モデル生物を用いる行動遺伝学者は、遺伝子を改変、挿入、または欠失させるために、様々な分子技術を用いる。これらの技術には、ノックアウト、フロキシング、遺伝子ノックダウン、CRISPR-Cas9などの方法を用いたゲノム編集などがある[19]。これらの技術により、行動遺伝学者は、モデル生物のゲノムにおいて異なるレベルの制御を可能にし、遺伝的変化の分子的、生理学的、または行動的な結果を評価することができる[20]。行動遺伝学でモデル生物としてよく用いられる動物には、マウス[21]、ゼブラフィッシュ[22]、線形動物のカエノラブディティス・エレガンスなどがある[23]。
機械学習と人工知能の発展により、研究者は複雑さと生成される大規模なデータセットを管理できる実験を設計できるようになり、ますます複雑な行動実験が可能になっている[24]。
ヒト研究
編集行動遺伝学研究で使用される研究デザインの一部は、双生児研究や養子研究を含む、家系デザイン(系図デザインとしても知られる)のバリエーションである[14]。遺伝関係が既知の個人(例えば、親子、兄弟、二卵性双生児、一卵性双生児)の量的遺伝学モデリングでは、個人間の表現型の違いに、遺伝子と環境がどの程度寄与しているかを推定することができる[25]。
双生児研究と家族研究
編集
双生児研究の基本的な直感は、一卵性双生児は100%のゲノムを共有し、二卵性双生児は平均して50%の分離ゲノムを共有するということである。したがって、一卵性双生児ペアの2人の間の違いは、環境の違いによってのみ生じる可能性があるのに対し、二卵性双生児は、環境だけでなく遺伝子によっても異なる。この単純化されたモデルでは、二卵性双生児の方が一卵性双生児よりも異なる場合、それは遺伝的影響にのみ帰することができる。双生児モデルの重要な仮定は、等しい環境の仮定であり[26]、一卵性双生児は二卵性双生児と同じ共有環境体験を持つというものである。例えば、一卵性双生児が二卵性双生児よりも似たような経験をする傾向があり、これらの経験自体が遺伝子-環境相関メカニズムを介して遺伝的に媒介されていない場合、一卵性双生児は二卵性双生児よりも互いに似ている傾向があり、その理由は遺伝子とは関係がない[27]。一卵性双生児と二卵性双生児の双生児研究では、双生児の類似性に影響を与える要因を記述し、遺伝率を推定するために、生物測定学的定式化を用いる[25][28]。この定式化は、表現型の分散が遺伝子と環境の2つの要因によるものであるという基本的な観察に基づいている。より正式には、 であり、ここで は表現型、 は遺伝子の影響、 は環境の影響、 は遺伝子と環境の相互作用である。 項は、相加的遺伝効果( )、優性効果( )、エピスタシス効果( )を含むように拡張できる。同様に、環境項 は、共有環境( )と非共有環境( )を含むように拡張できる。非共有環境には、測定誤差が含まれる。簡単のために遺伝子と環境の相互作用を除外し(双生児研究では典型的)、 項と 項を完全に分解すると、 となる。双生児研究では、この分解の単純化された形式を用いて、一卵性双生児と二卵性双生児の類似性をモデル化する(表参照)[25]。











| 関係の種類 | 完全分解 | ファルコナーの分解 |
|---|---|---|
| きょうだい間の完全な類似性 | ||
| 一卵性双生児の相関 ( ) | ||
| 二卵性双生児の相関 ( ) | ||
| ここで、 は未知の(おそらく非常に小さい)量である。 | ||









次に、単純化されたファルコナーの定式化を用いて、 、 、 の推定値を導出できる。 方程式と 方程式を並べ替えて代入することで、加算的遺伝分散、すなわち遺伝率の推定値 、非共有環境効果 、最後に共有環境効果 を得ることができる[25]。ここでは、双生児モデルがどのように機能するかを説明するために、ファルコナーの定式化を示した。現代のアプローチでは、最尤法を用いて、遺伝的および環境的な分散成分を推定している[29]。



ゲノムワイド関連解析
編集この節の加筆が望まれています。 |

行動遺伝学は、ゲノム情報、神経学的情報(神経解剖学/神経イメージングデータなど)、行動情報(ライフスタイルや健康に関連する活動に関する調査データなど)を含むデータベースを使用・生成する。このようなデータベースは、ゲノムワイド関連解析(GWAS)に使用される。
測定された遺伝的変異
編集ヒトゲノム計画により、科学者はDNAヌクレオチドの配列を直接ジェノタイピングできるようになった[32]。ジェノタイピングが行われると、遺伝的変異と精神障害、認知能力、性格などの行動表現型との関連性を検証できる[33]。
- 候補遺伝子 一つの人気のあるアプローチは、行動表現型との関連性について候補遺伝子を検証することであり、候補遺伝子は、行動形質や表現型の発現に関与する生物学的メカニズムに関する何らかの事前の理論に基づいて選択される[34]。一般に、このような研究は広く再現することが難しいことが証明されており[35][36][37][38]、このタイプの研究では偽陽性率が高いのではないかという懸念が示されている[34][39]。
- ゲノムワイド関連解析 ゲノムワイド関連解析では、研究者はゲノム全体にわたる数百万の遺伝的多型と行動表現型との関係を検証する[33]。この遺伝的関連解析へのアプローチは、表現型に関する特定の生物学的仮説に導かれることなく、大部分が理論に依存しない[33]。行動形質や精神疾患に関する遺伝的関連の知見は、多くの小さな遺伝的効果を含む高度に多因子的であることが分かっている[40][41][42][43][44]。
- SNP遺伝率と共遺伝率 最近、研究者は、一塩基多型(SNP)で測定された古典的に無関係な人々の間の類似性を用いて、ゲノムワイド複合形質解析(GCTA)などのソフトウェアに実装された混合効果モデルを用いて、SNPでタグ付けされた遺伝的変異や共変異を推定し始めている[45][46]。これを行うために、研究者は(通常は大きな)サンプル内のすべての個人間のすべてのSNPにおける平均的な遺伝的関連性を見出し、ヘイズマン-エルストン回帰または制限付き最尤法を用いて、SNPによって「タグ付け」または予測される遺伝的変異を推定する。表現型変異のうち、遺伝的関連性によって説明される割合は、「SNP遺伝率」と呼ばれている[47]。直感的には、測定されたSNPにおける遺伝的類似性によって表現型の類似性が予測される程度に応じて、SNP遺伝率は増加し、測定されたSNPが(典型的には稀な)原因となるバリアントをタグ付けできない程度に応じて、真の狭義の遺伝率よりも低くなると予想される[48]。この方法の価値は、双生児研究や家族研究と同じ仮定を必要としない遺伝率を推定する独立した方法であり、形質の変異の原因となるバリアントの対立遺伝子頻度スペクトルに関する洞察を与えることである[49]。
疑似実験デザイン
編集行動の遺伝的影響を理解するためではなく、行動に対する環境を介した影響を検証するために遺伝的影響を制御するために、いくつかの行動遺伝学的デザインは有用である[50]。このような行動遺伝学的デザインは、自然実験のサブセットと見なすことができる[51]。自然実験は、独立変数に対してある程度の制御を提供することで、真の実験を模倣する自然発生的な状況を利用しようとする疑似実験である。自然実験は、実際的または倫理的な制限から実験が実行不可能な場合に特に有用である[51]。
観察研究の一般的な限界は、遺伝子と環境の相対的影響が交絡していることである。この事実の単純な実証は、「環境」的影響の尺度が遺伝的であることである[52]。したがって、環境的リスク要因と健康アウトカムとの間に相関が観察されたとしても、必ずしも環境が健康アウトカムに影響を与えていることの証拠にはならない。同様に、例えば親子間の行動伝達に関する観察研究では、受動的な遺伝子-環境相関の問題により、伝達が遺伝的影響によるものか環境的影響によるものかを知ることは不可能である[51]。薬物を使用する親の子供が成人してから薬物を使用する可能性が高いという単純な観察では、子供が成長したときになぜ薬物を使用する可能性が高いのかを示すものではない。それは、子供が親の行動をモデリングしているからかもしれない。同様に、子供が親から薬物使用を促進する遺伝子を受け継いだため、親の行動に関係なく、成人してから薬物を使用するリスクが高くなったのかもしれない。養育環境と遺伝的継承の相対的な影響を解明する養子研究では、養子における喫煙、アルコール、マリファナの使用に対する養育環境の影響は小さいか無視できる程度であると分かっているが[53][要非一次資料]、ハードドラッグの使用に対する養育環境の影響はより大きいことが分かっている[54][要非一次資料]。
他の行動遺伝学デザインには、不一致双生児研究[50]、双生児の子供デザイン[55]、メンデルランダム化などがある[56]。
一般的な知見
編集行動の本質と起源に関して、行動遺伝学研究から得られる幅広い結論は多数ある[3][57]。3つの主要な結論は以下の通りである[3]。
- すべての行動形質と障害は遺伝子の影響を受ける
- 環境的影響は、同じ家族のメンバーを似通ったものにするのではなく、むしろ異なるものにする傾向がある
- 個人が年齢を重ねるにつれて、遺伝子の影響は相対的に重要性を増す傾向がある。
行動に対する遺伝的影響は広範である
編集研究されたすべての行動形質と障害が遺伝子の影響を受けている、つまり遺伝的であることは、複数の証拠から明らかである。最大の証拠源は双生児研究であり、一卵性(同一)双生児は同性の二卵性(二人の)双生児よりも互いに類似していることが常に観察されている[11][12]。
遺伝的影響が広範に及んでいるという結論は、双生児法の仮定に依存しない研究デザインでも観察されている。養子研究では、養子は様々な形質や障害について、養親よりも生物学的親族の方によく似ていることが常に示されている[3]。離れて育てられた双子のミネソタ研究では、生まれてすぐに離ればなれになった一卵性双生児が成人期に再会した[58]。これらの養子に出され、離れて育てられた双生児は、一般的な認知能力、人格、宗教的態度、職業的興味など、幅広い尺度で一緒に育てられた双生児と同じくらい似ていた[58]。ゲノムワイド遺伝子型判定を用いたアプローチにより、研究者は個人間の遺伝的関連性を測定し、何百万もの遺伝的変異に基づいて遺伝率を推定することができるようになった。名目上無関係な個人(近親者でも遠縁者でもない個人)間の遺伝的類似性(別名、関連性)の程度が表現型の類似性と関連しているかどうかを検証する方法が存在する[46]。このような方法は、双生児研究や養子研究と同じ仮定に依存しておらず、行動形質や障害の遺伝性の証拠を日常的に見出している[42][44][59]。
環境的影響の性質
編集研究された全てのヒトの行動表現型が遺伝子の影響を受けている(すなわち、遺伝的である)のと同様に、そのような表現型はすべて環境の影響も受けている[11][57]。一卵性双生児が遺伝的に同一でありながら、精神障害で完全に一致したり、行動形質で完全に相関したりすることは決してないという基本的な事実は、環境がヒトの行動を形作ることを示している[57]。
しかし、この環境的影響の性質は、同じ家族の個人をより似通ったものにするのではなく、互いにより異なったものにする傾向がある[3]。つまり、ヒトの研究における共有環境効果( )の推定値は、行動形質や精神障害の大多数で小さく、無視できるほど小さいか、ゼロであるのに対し、非共有環境効果( )の推定値は中程度から大きい[11]。双生児研究では、一卵性双生児間の相関( )が二卵性双生児の相関( )の少なくとも2倍あるため、 は通常0と推定される。ファルコナーの分散分解( )を使用すると、一卵性双生児と二卵性双生児の類似性のこの差は、 と推定されることになる。ファルコナーの分解は単純化されている[25]。優性効果とエピスタシス効果の影響を取り除いているが、これらの効果が存在する場合、一卵性双生児は二卵性双生児よりも似ている傾向があり、共有環境効果の影響を隠してしまう[25]。これは、 を推定するための双生児デザインの限界である。しかし、共有環境効果が無視できるという一般的な結論は、双生児研究だけに依存しているわけではない。養子研究でも、大きな( )要素は見出されていない。つまり、養親とその養子は、養子とその養育していない生物学的親よりもお互いに似ていない傾向がある[3]。少なくとも1人の実子と1人の養子がいる養子家族の研究では、研究されたほとんどの形質について、きょうだいの類似性もほぼゼロである傾向がある[11][60]。

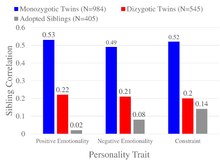
この図は、パーソナリティ研究の例であり、双生児研究と養子研究が、ポジティブ感情性、ネガティブ感情性、制約を含む多次元人格質問紙によって測定された幅広いパーソナリティ特性に対する共有環境の影響がゼロから小さいという結論に収束していることを示している[61]。
研究されたすべての行動形質と精神障害が遺伝的であるという結論が出ていることを考えると、生物学的きょうだいは、養子のきょうだいよりも常にお互いに似ている傾向があるだろう。しかし、特に思春期に測定されたいくつかの形質については、養子のきょうだいはお互いにある程度の有意な類似性(例えば、0.20の相関)を示す。共有環境の影響が有意であることが示されている形質には、内在化および外在化障害[62]、物質使用[63][要非一次資料] および依存症[54][要非一次資料]、 知能が含まれる[63][要非一次資料]。
遺伝的影響の性質
編集ヒトの行動結果に対する遺伝的影響は、複数の方法で記述できる[25]。その効果を記述する1つの方法は、行動の分散のうち、対立遺伝子が遺伝的変異でどの程度説明できるかという観点から、言い換えれば、決定係数または としても知られている。 について直感的に考える方法は、遺伝的変異が、異なる対立遺伝子を有する個人を、行動結果に関してどの程度互いに異なるようにさせるかを記述することである。個々の遺伝的変異の効果を記述する補完的な方法は、個人が有するリスク対立遺伝子の数の変化に対して、行動結果にどの程度の変化が期待されるかという点であり、多くの場合、ギリシャ文字の (回帰方程式の傾きを表す)で表されるか、二値の疾患結果の場合は、対立遺伝子の状態による疾患のオッズ比 で表される。違いに注意すること。 は、遺伝的変異内の対立遺伝子の集団レベルの効果を表す。 または は、リスク対立遺伝子を有さない個人と比較して、リスク対立遺伝子を有する個人に対する効果を表す[64]。



指標で記述された場合、複雑なヒトの行動形質や障害に対する個々の遺伝的変異の効果は極めて小さく、各変異は表現型の変動の を説明するに過ぎない[3]。この事実は、主に、物質使用[65][66]、人格[67]、出生力[68]、統合失調症[41]、うつ病[67][69]、脳の構造[70]や機能[71]などのエンドフェノタイプを含む複雑な行動表現型のゲノムワイド関連研究の結果を通して発見されてきた。この規則には、APOEのアルツハイマー病に対する効果[72]、CHRNA5の喫煙行動に対する効果[65]、(東アジア系の個人における)ALDH2のアルコール使用に対する効果など、少数の再現性があり頑健に研究された例外がある[73]。

一方、 指標に基づいて効果を評価した場合、複雑な行動表現型に非常に大きな効果を持つ遺伝的変異が多数存在する。そのような変異内のリスク対立遺伝子は極めてまれであり、その大きな行動的効果は少数の個人にのみ影響を与える。したがって、 指標を用いて集団レベルで評価した場合、集団内の個人間のリスクの差のうち、説明できるのはごくわずかである。例としては、APP内の変異があり、これは家族性の重度の早期発症型アルツハイマー病をもたらすが、比較的少数の個人にしか影響を与えない。これをAPOE内のリスク対立遺伝子と比較すると、APPと比べてリスクははるかに小さいが、はるかに一般的であるため、集団のより大きな割合に影響を与える[74]。
最後に、ハンチントン病のように、その病因が遺伝的に単純な古典的な行動障害がある。ハンチントン病は、HTT遺伝子の単一の常染色体優性変異によって引き起こされ、この変異は、個人が十分に長生きすると仮定した場合、この病気を発症するリスクの個人差を説明する唯一の変異である[75]。ハンチントン病のような遺伝的に単純でまれな疾患の場合、変異の と は同時に大きくなる[64]。
その他の一般的な知見
編集心理学研究の再現性に関する一般的な懸念に対応して、行動遺伝学者のロバート・プロミン、ジョン・C・デフリーズ、バレリー・クノピック、ジェナエ・ナイダーハイザーは、行動遺伝学研究から得られた最も再現性の高い10の知見のレビューを発表した[57]。その10の知見は以下の通りである。
- すべての心理的形質は、有意かつ実質的な遺伝的影響を示す。
- 100%遺伝する行動形質はない。
- 遺伝率は、小さな効果を持つ多くの遺伝子によって引き起こされる。
- 心理的形質間の表現型相関は、有意かつ実質的な遺伝的媒介を示す。
- 知能の遺伝率は、発達を通して増加する。
- 年齢間の安定性は、主に遺伝によるものである。
- 「環境」の尺度のほとんどは、有意な遺伝的影響を示す。
- 環境的尺度と心理的形質の間のほとんどの関連は、遺伝的に有意に媒介されている。
- ほとんどの環境的効果は、同じ家族で育つ子供によって共有されない。
- 異常は正常である。
批判と論争
編集行動遺伝学の研究と知見は、時に論争を引き起こしてきた。この論争の一部は、行動遺伝学の知見が、人間の行動と能力の本質に関する社会的信念に異議を唱える可能性があるために生じたものである。主要な論争の分野には、人種差、知能、暴力、人間の性などのトピックに関する遺伝学的研究が含まれる[76]。その他の論争は、一般の人々や研究者自身による行動遺伝学研究の誤解から生じている[3]。例えば、遺伝率の概念は、因果関係を意味するものだと誤解されやすく、ある行動や状態が遺伝的素質によって決定されるという意味だと誤解されやすい[77]。行動遺伝学の研究者が、ある行動がX%遺伝すると言った場合、それは遺伝学がその行動の最大X%を引き起こしたり、決定したり、修正したりすることを意味するのではない。むしろ、遺伝率は、集団レベルで形質の差異と相関する遺伝的差異に関する記述である[要出典]。
歴史的に見て、おそらく最も論争の的となった主題は、人種と遺伝学に関するものであった[76]。人種は科学的に正確な用語ではなく、その解釈は文化や出身国によって異なる場合がある[78]。その代わりに、遺伝学者は祖先などのより厳密に定義された概念を使用する[79]。例えば、いわゆる「黒人」の人種には、比較的最近のアフリカ系の個人がすべて含まれる可能性がある(すべての人類がアフリカの祖先から進化したので、「最近」という意味である)。しかし、アフリカには世界の他の地域を合わせたよりも多くの遺伝的多様性がある[80]。そのため、"黒人"という人種を語ることは、正確な遺伝学的意味を持たない[79]。
定性的研究では、行動遺伝学が科学的規範やコンセンサスのない統制不能な分野であり、それが論争を助長しているという議論が促進されてきた。この議論は、このような状況が、人種、知能、単一の遺伝子内の変異が論争の的となる表現型に非常に強く影響することが発見された事例(例えば、「ゲイ遺伝子」論争)などの論争につながったと続けている。この議論は、さらに、行動遺伝学における論争の持続と、論争の解決の失敗のために、行動遺伝学は優れた科学の基準に適合していないと述べている[81]。
行動遺伝学研究の一部の基礎となる科学的仮定も、欠陥があると批判されている[77]。ゲノムワイド関連解析では、相加性など、統計的には頑健だが一部の行動については非現実的な単純化した統計的仮定を用いることが多い。批判者は、さらに、ヒトにおいて、行動遺伝学は統計解析の不正確な解釈に基づく遺伝的還元主義の誤った形態を表していると主張する[82]。一卵性(MZ)双生児と二卵性(DZ)双生児を比較した研究では、環境の影響は両方の双生児で同じになると仮定しているが、この仮定も非現実的かもしれない。一卵性双生児は二卵性双生児よりも似たような扱いを受ける可能性があり[77]、これ自体が、自分の遺伝子が他人からの扱いに影響を与えるという、喚起的な遺伝子-環境相関の一例である可能性がある。また、双生児研究では、共有された子宮内環境の影響を排除することはできないが、一絨毛膜と二絨毛膜の子宮内環境を経験する双生児を比較する研究は存在し、限定的な影響を示唆している[83]。生後早期に離ればなれになった双生児の研究には、生まれてすぐにではなく、幼児期の途中で離ればなれになった子供が含まれる[77]。したがって、そのような研究では、早期に離ればなれになった双生児と後に離ればなれになった双生児の類似性を比較することで、ある程度、早期の養育環境の影響を評価することができる[58]。
出典
編集- ^ a b “History of Behavior Genetics”. Handbook of Behavior Genetics (1 ed.). New York, NY: Springer. (2009). pp. 3–11. doi:10.1007/978-0-387-76727-7_1. ISBN 978-0-387-76726-0
- ^ a b “A History of Behavior Genetics”. Neurobehavioral Genetics: Methods and Applications, Second Edition. CRC Press. (30 August 2006). ISBN 978-1-4200-0356-7
- ^ a b c d e f g h i j k l m n o McGue M, Gottesman II (2015). "Behavior Genetics". The Encyclopedia of Clinical Psychology. pp. 1–11. doi:10.1002/9781118625392.wbecp578. ISBN 9781118625392。
- ^ The Tempest. The Arden Shakespeare (Third ed.). The Arden Shakespeare. (1999). p. 60. ISBN 978-1-903436-08-0
- ^ Hereditary Genius: An Inquiry into Its Laws and Consequences. London: MacMillan and Co.. (1869). オリジナルの7 December 2019時点におけるアーカイブ。 17 December 2009閲覧。
- ^ “Darwin, Galton and the Statistical Enlightenment”. Journal of the Royal Statistical Society, Series A 173 (3): 469–482. (July 2010). doi:10.1111/j.1467-985X.2010.00643.x.
- ^ Hall CS (1951). “The genetics of behavior”. In Stevens SS. Handbook of Experimental Psychology. New York: John Wiley and Sons. pp. 304–329
- ^ “Russian psychogenetics”. Psychology of Russia: Past, Present, Future. Commack, NY: Nova Science. (1997). pp. 83–124
- ^ “Psychogenetics of emotionality in the rat”. Annals of the New York Academy of Sciences 159 (3): 806–24. (July 1969). Bibcode: 1969NYASA.159..806B. doi:10.1111/j.1749-6632.1969.tb12980.x. PMID 5260300.
- ^ Behavior Genetics. New York: John Wiley and Sons. (1960)
- ^ a b c d e “Meta-analysis of the heritability of human traits based on fifty years of twin studies”. Nature Genetics 47 (7): 702–9. (July 2015). doi:10.1038/ng.3285. PMID 25985137. オリジナルの2022-04-20時点におけるアーカイブ。 2019年1月30日閲覧。.
- ^ a b “Three Laws of Behavior Genetics and What They Mean”. Current Directions in Psychological Science 9 (5): 160–164. (2000). doi:10.1111/1467-8721.00084. オリジナルの2016-10-09時点におけるアーカイブ。 2016年4月12日閲覧。.
- ^ “Publication Trends Over 55 Years of Behavioral Genetic Research”. Behavior Genetics 46 (5): 603–7. (September 2016). doi:10.1007/s10519-016-9786-2. PMC 5206393. PMID 26992731.
- ^ a b Behavioral Genetics. Worth Publishers. (24 September 2012). ISBN 978-1-4292-4215-8. オリジナルの31 December 2013時点におけるアーカイブ。 27 January 2016閲覧。
- ^ “behavior genetics | Definition, History, & Methods | Britannica”. web.archive.org (2021年4月17日). 2024年3月11日閲覧。
- ^ “Response to 20 generations of selection for open-field activity in mice”. Behavioral Biology 11 (4): 481–95. (August 1974). doi:10.1016/s0091-6773(74)90800-1. PMID 4415597.
- ^ “Response to divergent selection for nesting behavior in Mus musculus”. Genetics 96 (3): 757–65. (November 1980). doi:10.1093/genetics/96.3.757. PMC 1214374. PMID 7196362.
- ^ “Artificial selection for increased wheel-running behavior in house mice”. Behavior Genetics 28 (3): 227–37. (May 1998). doi:10.1023/A:1021479331779. PMID 9670598.
- ^ “Applications of CRISPR-Cas systems in neuroscience”. Nature Reviews. Neuroscience 17 (1): 36–44. (January 2016). doi:10.1038/nrn.2015.2. PMC 4899966. PMID 26656253.
- ^ “A mouse geneticist's practical guide to CRISPR applications”. Genetics 199 (1): 1–15. (January 2015). doi:10.1534/genetics.114.169771. PMC 4286675. PMID 25271304.
- ^ “The ascent of mouse: advances in modelling human depression and anxiety” (英語). Nature Reviews. Drug Discovery 4 (9): 775–790. (September 2005). doi:10.1038/nrd1825. PMID 16138108.
- ^ “Behavioral genetics in larval zebrafish: learning from the young”. Developmental Neurobiology 72 (3): 366–372. (March 2012). doi:10.1002/dneu.20872. PMC 6430578. PMID 22328273.
- ^ “The behavioral genetics of Caenorhabditis elegans”. Behavior Genetics 20 (2): 169–189. (March 1990). doi:10.1007/bf01067789. PMID 2191646.
- ^ “Complex Economic Behavior Patterns Are Constructed from Finite, Genetically Controlled Modules of Behavior”. Cell Reports 28 (7): 1814–1829.e6. (August 2019). doi:10.1016/j.celrep.2019.07.038. PMC 7476553. PMID 31412249.
- ^ a b c d e f g h Introduction to quantitative genetics. Longman, Scientific & Technical. (1989). ISBN 978-0-470-21162-5. オリジナルの2021-04-22時点におけるアーカイブ。 2016年12月2日閲覧。
- ^ “Has the "Equal Environments" assumption been tested in twin studies?”. Twin Research 6 (6): 486–9. (2003). doi:10.1375/136905203322686473. PMID 14965458.
- ^ “A test of the equal-environment assumption in twin studies of psychiatric illness”. Behavior Genetics 23 (1): 21–7. (January 1993). doi:10.1007/BF01067551. PMID 8476388.
- ^ “Comparison of the biometrical genetical, MAVA, and classical approaches to the analysis of the human behavior.”. Psychological Bulletin 73 (5): 311–349. (1970). doi:10.1037/h0029135. PMID 5528333.
- ^ “The genetical analysis of covariance structure”. Heredity 38 (1): 79–95. (February 1977). doi:10.1038/hdy.1977.9. PMID 268313.
- ^ “Massive genome study informs the biology of reading and language” (英語). Max Planck Society via medicalxpress.com 18 September 2022閲覧。
- ^ “Genome-wide analyses of individual differences in quantitatively assessed reading- and language-related skills in up to 34,000 people”. Proceedings of the National Academy of Sciences of the United States of America 119 (35): e2202764119. (August 2022). Bibcode: 2022PNAS..11902764E. doi:10.1073/pnas.2202764119. PMC 9436320. PMID 35998220.
- ^ “Initial impact of the sequencing of the human genome”. Nature 470 (7333): 187–97. (February 2011). Bibcode: 2011Natur.470..187L. doi:10.1038/nature09792. hdl:1721.1/69154. PMID 21307931.
- ^ a b c “Genome-wide association studies for complex traits: consensus, uncertainty and challenges”. Nature Reviews Genetics 9 (5): 356–69. (May 2008). doi:10.1038/nrg2344. PMID 18398418.
- ^ a b “A critical review of the first 10 years of candidate gene-by-environment interaction research in psychiatry”. The American Journal of Psychiatry 168 (10): 1041–9. (October 2011). doi:10.1176/appi.ajp.2011.11020191. PMC 3222234. PMID 21890791.
- ^ “Evaluating historical candidate genes for schizophrenia”. Molecular Psychiatry 20 (5): 555–562. (May 2015). doi:10.1038/mp.2015.16. PMC 4414705. PMID 25754081.
- ^ “Editorial policy on candidate gene association and candidate gene-by-environment interaction studies of complex traits”. Behavior Genetics 42 (1): 1–2. (January 2012). doi:10.1007/s10519-011-9504-z. PMID 21928046.
- ^ “No Evidence That Schizophrenia Candidate Genes Are More Associated With Schizophrenia Than Noncandidate Genes”. Biological Psychiatry. Risk Genes and the Emergence of Schizophrenia 82 (10): 702–708. (November 2017). doi:10.1016/j.biopsych.2017.06.033. PMC 5643230. PMID 28823710.
- ^ “No Support for Historical Candidate Gene or Candidate Gene-by-Interaction Hypotheses for Major Depression Across Multiple Large Samples”. The American Journal of Psychiatry 176 (5): 376–387. (May 2019). doi:10.1176/appi.ajp.2018.18070881. PMC 6548317. PMID 30845820.
- ^ “Problems of reporting genetic associations with complex outcomes”. Lancet 361 (9360): 865–872. (March 2003). doi:10.1016/S0140-6736(03)12715-8. PMID 12642066.
- ^ “Five years of GWAS discovery”. American Journal of Human Genetics 90 (1): 7–24. (January 2012). doi:10.1016/j.ajhg.2011.11.029. PMC 3257326. PMID 22243964.
- ^ a b “Biological insights from 108 schizophrenia-associated genetic loci”. Nature 511 (7510): 421–7. (July 2014). Bibcode: 2014Natur.511..421S. doi:10.1038/nature13595. PMC 4112379. PMID 25056061.
- ^ a b “Estimating the proportion of variation in susceptibility to schizophrenia captured by common SNPs”. Nature Genetics 44 (3): 247–50. (February 2012). doi:10.1038/ng.1108. PMC 3327879. PMID 22344220.
- ^ “Genetic architectures of psychiatric disorders: the emerging picture and its implications”. Nature Reviews. Genetics 13 (8): 537–51. (July 2012). doi:10.1038/nrg3240. PMC 4110909. PMID 22777127.
- ^ a b “Meta-analysis of Genome-wide Association Studies for Neuroticism, and the Polygenic Association With Major Depressive Disorder”. JAMA Psychiatry 72 (7): 642–50. (July 2015). doi:10.1001/jamapsychiatry.2015.0554. PMC 4667957. PMID 25993607.
- ^ “Common SNPs explain a large proportion of the heritability for human height”. Nature Genetics 42 (7): 565–9. (July 2010). doi:10.1038/ng.608. PMC 3232052. PMID 20562875.
- ^ a b “GCTA: a tool for genome-wide complex trait analysis”. American Journal of Human Genetics 88 (1): 76–82. (January 2011). doi:10.1016/j.ajhg.2010.11.011. PMC 3014363. PMID 21167468.
- ^ “Estimation of SNP heritability from dense genotype data”. American Journal of Human Genetics 93 (6): 1151–5. (2013). doi:10.1016/j.ajhg.2013.10.015. PMC 3852919. PMID 24314550.
- ^ “A commentary on 'common SNPs explain a large proportion of the heritability for human height' by Yang et al. (2010)”. Twin Research and Human Genetics 13 (6): 517–24. (2010). doi:10.1375/twin.13.6.517. PMID 21142928.
- ^ “Research review: Polygenic methods and their application to psychiatric traits”. Journal of Child Psychology and Psychiatry, and Allied Disciplines 55 (10): 1068–87. (2014). doi:10.1111/jcpp.12295. PMID 25132410. オリジナルの2017-01-16時点におけるアーカイブ。 2019年7月1日閲覧。.
- ^ a b “Causal Inference and Observational Research: The Utility of Twins”. Perspectives on Psychological Science 5 (5): 546–56. (September 2010). doi:10.1177/1745691610383511. PMC 3094752. PMID 21593989.
- ^ a b c “Proceeding From Observed Correlation to Causal Inference: The Use of Natural Experiments”. Perspectives on Psychological Science 2 (4): 377–95. (December 2007). doi:10.1111/j.1745-6916.2007.00050.x. PMID 26151974.
- ^ “Genetic influences on measures of the environment: a systematic review”. Psychological Medicine 37 (5): 615–26. (May 2007). doi:10.1017/S0033291706009524. PMID 17176502.
- ^ “Parental smoking and adolescent problem behavior: an adoption study of general and specific effects”. The American Journal of Psychiatry 165 (10): 1338–44. (October 2008). doi:10.1176/appi.ajp.2008.08010125. PMC 2597022. PMID 18676589.
- ^ a b “Genetic and familial environmental influences on the risk for drug abuse: a national Swedish adoption study”. Archives of General Psychiatry 69 (7): 690–7. (July 2012). doi:10.1001/archgenpsychiatry.2011.2112. PMC 3556483. PMID 22393206.
- ^ “The role of the children of twins design in elucidating causal relations between parent characteristics and child outcomes”. Journal of Child Psychology and Psychiatry, and Allied Disciplines 44 (8): 1130–44. (November 2003). doi:10.1111/1469-7610.00196. PMID 14626455.
- ^ “Mendelian randomization: prospects, potentials, and limitations”. International Journal of Epidemiology 33 (1): 30–42. (February 2004). doi:10.1093/ije/dyh132. PMID 15075143.
- ^ a b c d “Top 10 Replicated Findings From Behavioral Genetics”. Perspectives on Psychological Science 11 (1): 3–23. (January 2016). 27 January 2016. doi:10.1177/1745691615617439. PMC 4739500. PMID 26817721.
- ^ a b c “Sources of human psychological differences: the Minnesota Study of Twins Reared Apart”. Science 250 (4978): 223–8. (October 1990). Bibcode: 1990Sci...250..223B. doi:10.1126/science.2218526. PMID 2218526.
- ^ “Common DNA markers can account for more than half of the genetic influence on cognitive abilities”. Psychological Science 24 (4): 562–8. (April 2013). doi:10.1177/0956797612457952. PMC 3652710. PMID 23501967.
- ^ “Why are children in the same family so different from one another?”. International Journal of Epidemiology 40 (3): 563–82. (June 2011). doi:10.1093/ije/dyq148. PMC 3147063. PMID 21807642.
- ^ “Shared environmental influences on personality: a combined twin and adoption approach”. Behavior Genetics 43 (6): 491–504. (November 2013). doi:10.1007/s10519-013-9616-8. PMC 3868213. PMID 24065564.
- ^ “Rethinking environmental contributions to child and adolescent psychopathology: a meta-analysis of shared environmental influences”. Psychological Bulletin 135 (4): 608–37. (July 2009). doi:10.1037/a0015702. PMID 19586164.
- ^ a b “Are there shared environmental influences on adolescent behavior? Evidence from a study of adoptive siblings”. Behavior Genetics 39 (5): 532–40. (September 2009). doi:10.1007/s10519-009-9283-y. PMC 2858574. PMID 19626434.
- ^ a b “Statistics notes. The odds ratio”. BMJ 320 (7247): 1468. (May 2000). doi:10.1136/bmj.320.7247.1468. PMC 1127651. PMID 10827061.
- ^ a b “Sequence variants at CHRNB3-CHRNA6 and CYP2A6 affect smoking behavior”. Nature Genetics 42 (5): 448–53. (May 2010). doi:10.1038/ng.573. PMC 3080600. PMID 20418888.
- ^ “Genome-wide association and genetic functional studies identify autism susceptibility candidate 2 gene (AUTS2) in the regulation of alcohol consumption”. Proceedings of the National Academy of Sciences of the United States of America 108 (17): 7119–24. (April 2011). Bibcode: 2011PNAS..108.7119S. doi:10.1073/pnas.1017288108. PMC 3084048. PMID 21471458.
- ^ a b “Genetic variants associated with subjective well-being, depressive symptoms, and neuroticism identified through genome-wide analyses”. Nature Genetics 48 (6): 624–33. (June 2016). doi:10.1038/ng.3552. PMC 4884152. PMID 27089181.
- ^ “Physical and neurobehavioral determinants of reproductive onset and success”. Nature Genetics 48 (6): 617–23. (June 2016). doi:10.1038/ng.3551. PMC 5238953. PMID 27089180.
- ^ “Sparse whole-genome sequencing identifies two loci for major depressive disorder”. Nature 523 (7562): 588–91. (July 2015). Bibcode: 2015Natur.523..588C. doi:10.1038/nature14659. PMC 4522619. PMID 26176920.
- ^ “Common genetic variants influence human subcortical brain structures”. Nature 520 (7546): 224–9. (April 2015). Bibcode: 2015Natur.520..224.. doi:10.1038/nature14101. PMC 4393366. PMID 25607358.
- ^ “Knowns and unknowns for psychophysiological endophenotypes: integration and response to commentaries”. Psychophysiology 51 (12): 1339–47. (December 2014). doi:10.1111/psyp.12358. PMC 4231488. PMID 25387720.
- ^ “Protective effect of apolipoprotein E type 2 allele for late onset Alzheimer disease”. Nature Genetics 7 (2): 180–4. (June 1994). doi:10.1038/ng0694-180. PMID 7920638.
- ^ “Meta-analyses of ALDH2 and ADH1B with alcohol dependence in Asians”. Psychological Bulletin 132 (4): 607–21. (July 2006). doi:10.1037/0033-2909.132.4.607. PMID 16822169.
- ^ “The genetic architecture of Alzheimer's disease: beyond APP, PSENs and APOE”. Neurobiology of Aging 33 (3): 437–56. (March 2012). doi:10.1016/j.neurobiolaging.2010.03.025. PMC 2980860. PMID 20594621.
- ^ “A polymorphic DNA marker genetically linked to Huntington's disease”. Nature 306 (5940): 234–8. (1983). Bibcode: 1983Natur.306..234G. doi:10.1038/306234a0. PMID 6316146.
- ^ a b “Ethics: Taboo genetics”. Nature 502 (7469): 26–8. (October 2013). Bibcode: 2013Natur.502...26C. doi:10.1038/502026a. PMID 24091964.
- ^ a b c d “Genes, behavior, and behavior genetics”. Wiley Interdisciplinary Reviews: Cognitive Science 8 (1–2): e1405. (January 2017). doi:10.1002/wcs.1405. hdl:10161/13337. PMID 27906529.
- ^ “Science and Society: Taking race out of human genetics”. Science 351 (6273): 564–5. (February 2016). Bibcode: 2016Sci...351..564Y. doi:10.1126/science.aac4951. PMID 26912690.
- ^ a b “The genetic ancestry of African Americans, Latinos, and European Americans across the United States”. American Journal of Human Genetics 96 (1): 37–53. (January 2015). doi:10.1016/j.ajhg.2014.11.010. PMC 4289685. PMID 25529636.
- ^ “An integrated map of genetic variation from 1,092 human genomes”. Nature 491 (7422): 56–65. (November 2012). Bibcode: 2012Natur.491...56T. doi:10.1038/nature11632. PMC 3498066. PMID 23128226.
- ^ Misbehaving Science: Controversy and the Development of Behavior Genetics. University of Chicago Press. (7 July 2014). ISBN 978-0-226-05859-7. オリジナルの22 April 2021時点におけるアーカイブ。 23 May 2018閲覧。
- ^ “Eliminating Genetic Reductionism from Developmental Science”. Research in Human Development 12 (3–4): 178–188. (27 August 2015). doi:10.1080/15427609.2015.1068058. ISSN 1542-7609.
- ^ “Chorionicity and Heritability Estimates from Twin Studies: The Prenatal Environment of Twins and Their Resemblance Across a Large Number of Traits”. Behavior Genetics 46 (3): 304–14. (May 2016). doi:10.1007/s10519-015-9745-3. PMC 4858554. PMID 26410687.
参考文献
編集- “Key issues in contemporary behavioral genetics”. Current Opinion in Behavioral Sciences 2: 89–95. (2015). doi:10.1016/j.cobeha.2014.10.002.
- Handbook of Molecular-Genetic Techniques for Brain and Behavior Research. Techniques in the Behavioral and Neural Sciences. 13. Elsevier. (1999). ISBN 978-0-444-50239-1
- Behavioral Genetics of the Mouse: Genetics of Behavioral Phenotypes. Cambridge Handbooks in Behavioral Genetics. 1. Cambridge University Press. (2013). ISBN 978-1-107-03481-5
- How Genes Influence Behavior. Oxford University Press. (28 January 2010). ISBN 978-0-19-955990-9
- “Beyond Heritability: Twin Studies in Behavioral Research”. Current Directions in Psychological Science 18 (4): 217–220. (August 2010). doi:10.1111/j.1467-8721.2009.01639.x. PMC 2899491. PMID 20625474.
- “Understanding Heritability: What it is and What it is Not”. European Journal of Personality 25 (4): 287–294. (2011). doi:10.1002/per.835. ISSN 0890-2070. オリジナルの16 December 2013時点におけるアーカイブ。 15 December 2013閲覧。.
- “Chapter 1: Behavioral Genetics”. Handbook of Psychology (Volume 3: Behavioral Neuroscience). John Wiley & Sons. (10 October 2012). ISBN 978-0-470-89059-2. オリジナルの16 December 2013時点におけるアーカイブ。 15 December 2013閲覧。
- “Chapter 10: Behavior Genetics”. The Wiley-Blackwell Handbook of Individual Differences. United Kingdom: Blackwell Publishing Ltd. (2011). doi:10.1002/9781444343120. ISBN 978-1-4443-3438-8
関連項目
編集外部リンク
編集- “Introduction to Human Behavioral Genetics”. Coursera (5 May 2014). 10 June 2014閲覧。
- “Introduction to Behavioral Genetics”. University of Minnesota (December 2020). 28 June 2021閲覧。
- “Behavior Genetics Association”. Behavior Genetics Association. 8 May 2022閲覧。
- Institute for Behavioral Genetics at the University of Colorado Boulder, University of Colorado Boulder
- “Virginia Institute for Psychiatric and Behavioral Genetics”. Virginia Commonwealth University. 2024年3月12日閲覧。