滑距目
滑距目(かっきょもく、Litopterna[1])とは、かつて南アメリカから北アメリカにかけて生息していた絶滅した蹄を持った動物のこと。
| 滑距目 Litopterna | |||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
.jpg) マクラウケニア(Macrauchenia)の想像図
| |||||||||||||||||||||||||||
| 保全状況評価 | |||||||||||||||||||||||||||
| 絶滅(化石) | |||||||||||||||||||||||||||
| 地質時代 | |||||||||||||||||||||||||||
| 暁新世 - 更新世 | |||||||||||||||||||||||||||
| 分類 | |||||||||||||||||||||||||||
|
進化史
編集最古の化石記録は暁新世の南アメリカから発見されていて、南蹄目ほど多様化していなかったが、南アメリカで繁栄した。この動物群の祖先は髁節目を祖先にした誕生したと考えられている。滑距目は他の大陸の有蹄類に類似する進化傾向が大きく分けて二つある。それはプロテロテリウム科とマクラウケニア科で、前者はウマ形亜目と類似していて、蹄が少なくなっていき、体はウマ形亜目に比べて大型化はしなかったが、疾走に適した体になっていた[2][3]。トアテリウムは蹄は一つにもなって、ウマ形亜目よりか早く指を一本にした[2][3][4]。また、奇蹄目の進化傾向の少臼歯の大臼歯化なども、類似している。中新世から鮮新世が進化のピークで[3]、鮮新世に絶滅した[2][3]。
後者は現在の進化の傾向は他の有蹄類と類似する点はすくないが、形態的には似ている点は多い、進化した種ほど外鼻孔が後退し、鼻が長かったと考えられる[5]。更新世に絶滅した[5][2]。
他の科のプロトリプテルナ科は、暁新世に出現し、始新世の移り変わりの時期には絶滅し、アディアントゥス科は中新世中期に絶滅した[2]。
形態
編集
初期の種でも肩高50cmで大きな種でも肩高1.6mで、進化史の節と同じで、大きく二つの形態に分けれる。一つは、プロテロテリウム科で、ウマ形亜目に類似していて、小柄であったが、指は第3指で体重を支えているが、第2指と第4指を残した種もいれば、完全に第3指のみになった種もいる[4][6]。また、頭骨の顔面部は長く低くなっていて、背中はまっすぐ、四肢は長くなっていて、脚の中軸が第3指を通るようになっており、距骨の上部の関節は滑車のようになっていて、これもウマ形亜目ににていた[6]。

二つ目は、マクラウケニア科で、全体的にラクダに似ており、前後に長い頭蓋と長い四肢と頸椎を持っていた[6]。3本指で、外鼻孔は後退し、ヘラジカに似た[7]発達した鼻を持っていたと考えられている[6]。また、歯は哺乳類の基本の歯の数、44本全てそろえており、他の科と比べて比較的に歯冠が高く、隙間があまりなかった[6][2]。
分布
編集暁新世に南アメリカで誕生し、長らく南アメリカで多様化したが、パナマ地峡が鮮新世に登場し、北アメリカに分布した[5][6]。
分類
編集上位系統
編集滑距目に属すマクラウケニアは、ミトコンドリアDNAの分析から、奇蹄目と姉妹群を成すことが明らかになった[8][9]。コラーゲンの分析でも、マクラウケニアは南蹄目のトクソドンとともに、奇蹄目と姉妹群をなすことが明らかになっている[10][11]。これにより、「汎奇蹄類」(Panperissodactyla)に含める分類が提唱されている[10]。
下位分類
編集以下、遠藤秀紀(2002)に従う[12]。
- 滑距目 Litopterna
系統
編集
ギャラリー
編集-
 トアテリウムの頭骨。
トアテリウムの頭骨。 -
 トアテリウムの全身骨格復元図。
トアテリウムの全身骨格復元図。 -
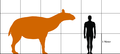 マクラウケニアと人間の大きさの比較。
マクラウケニアと人間の大きさの比較。 -
 トアテリウムの全身骨格。
トアテリウムの全身骨格。 -
 マクラウケニアの予想図。
マクラウケニアの予想図。

_(14749873255).jpg)

_and_Macrauchenia_patachonica_(far).jpg)
.jpg)
参考文献
編集- McKenna, M. C, and S. K. Bell. 1997. Classification of Mammals Above the Species Level. Columbia University Press, New York, 631 pp.
関連項目
編集外部リンク
編集An artist's rendition of a Macrauchenia, a representative genus of the Litopterna. Retrieved from the Red Académica Uruguaya megafauna page.
{kind=link}
脚注
編集- ^ 遠藤秀紀、佐々木基樹「哺乳類分類における高次群の和名について」『日本野生動物医学会誌』第6巻第2号、日本野生動物医学会、2001年、45-53頁、CRID 1390282679465041024、doi:10.5686/jjzwm.6.45、ISSN 1342-6133。
- ^ a b c d e f 『新版絶滅哺乳類図鑑』丸善、228頁。
- ^ a b c d 『脊椎動物の進化』築地書館、444頁。
- ^ a b 『古第三紀・新第三紀・第四紀』技術評論社。
- ^ a b c 『新版絶滅哺乳類図鑑』丸善、229頁。
- ^ a b c d e f 『脊椎動物の進化』築地書館、444,445頁。
- ^ Moyano, Silvana; Giannini, Norberto (2018-10-29). “Cranial characters associated with the proboscis postnatal-development in Tapirus (Perissodactyla: Tapiridae) and comparisons with other extant and fossil hoofed mammals”. Zoologischer Anzeiger - A Journal of Comparative Zoology 277: 143–147. doi:10.1016/j.jcz.2018.08.005.
- ^ Westbury, M.; Baleka, S.; Barlow, A.; Hartmann, S.; Paijmans, J. L. A.; Kramarz, A.; Forasiepi, A. M.; Bond, M. et al. (2017-06-27). “A mitogenomic timetree for Darwin's enigmatic South American mammal Macrauchenia patachonica”. Nature Communications 8: 15951. Bibcode: 2017NatCo...815951W. doi:10.1038/ncomms15951. PMC 5490259. PMID 28654082.
- ^ Strickland, A. (June 27, 2017). “DNA solves ancient animal riddle that Darwin couldn't”. CNN June 27, 2017閲覧。
- ^ a b Welker, F.; Collins, M. J.; Thomas, J. A.; Wadsley, M.; Brace, S.; Cappellini, E.; Turvey, S. T.; Reguero, M. et al. (2015-03-18). “Ancient proteins resolve the evolutionary history of Darwin's South American ungulates”. Nature 522 (7554): 81–84. Bibcode: 2015Natur.522...81W. doi:10.1038/nature14249. ISSN 0028-0836. PMID 25799987.
- ^ Buckley, M. (2015-04-01). “Ancient collagen reveals evolutionary history of the endemic South American 'ungulates'”. Proceedings of the Royal Society B: Biological Sciences 282 (1806): 20142671. doi:10.1098/rspb.2014.2671. PMC 4426609. PMID 25833851.
- ^ 『哺乳類の進化』東京大学出版会。
この項目は、古生物学に関連した書きかけの項目です。この項目を加筆・訂正などしてくださる協力者を求めています(プロジェクト:地球科学、プロジェクト:生物/Portal:地球科学、Portal:生物学)。 |