感覚ニューロン
感覚ニューロン(かんかくニューロン、英: sensory neuron)は、求心性ニューロン(きゅうしんせいニューロン、英: afferent neuron)とも呼ばれ、受容体を介して特定の種類の刺激を活動電位または段階的電位に変換する神経系のニューロン(神経細胞)である[1]。
このプロセスは、感覚伝達(sensory transduction)と呼ばれる。感覚ニューロンの細胞体は、脊髄の背根神経節にある[2]。
感覚情報は、感覚神経の求心性神経線維を通り、脊髄を経由して脳へと到達する。それらの刺激は、光や音など体外を感知するような外受容器から来る場合と、血圧や体位感覚に反応するような内受容器から来る場合とがある。
種類と機能
編集_J.PNG)
感覚ニューロンの種類によって、異なる種類の刺激に反応する異なる感覚受容器(sensory receptors)を持っている。少なくとも、6つの外部感覚受容器と2つの内部感覚受容器が知られている。
外受容器
編集体外からの刺激に反応する外部受容器を外受容器(exteroreceptors)と呼ぶ[3]。外受容器には、嗅覚受容体(匂い)、味覚受容体、光受容器(視覚)、有毛細胞(音)、温度受容器(熱さ寒さ)、そしてさまざまな機械受容器(伸縮、歪み)がある。
嗅覚
編集嗅覚に関与する感覚ニューロンは、嗅覚受容神経と呼ばれている。これらのニューロンには、嗅覚受容体と呼ばれる受容体があって、空気中の匂い分子によって活性化される。空気中の分子は、肥大した繊毛と微絨毛によって検出される[4]。これらの感覚ニューロンは活動電位を発生させる。その軸索は嗅神経を形成し、大脳皮質のニューロン(嗅球)に直接接合する。すなわち、他の感覚系と同じルートを使わず、脳幹や視床を迂回している。嗅球のニューロンは、感覚神経の入力を直接受けて、嗅覚系の他の部分や大脳辺縁系の多くの部分と結合している。
味覚
編集味蕾にある味覚受容器は、嗅覚受容体と同様に、食物中の化学物質と相互作用して活動電位を発生させる。
視覚
編集光受容細胞(視細胞)は、光(電磁放射)を電気信号に変換する光伝達(phototransduction)という機能を持っている。この信号は、網膜に存在する別種のニューロンとの相互作用によって精緻化され、制御される。網膜内には5種類の基本的なニューロンがあり、それぞれ光受容細胞、双極細胞、神経節細胞、水平細胞、アマクリン細胞という。網膜の基本的な回路には、光受容細胞(桿体または錐体のいずれか)、双極細胞、神経節細胞という3つのニューロンからなる連鎖が組み込まれている。最初の活動電位は、網膜神経節細胞で発生する。この経路は、視覚情報を脳に伝達する最も直接的な方法である。光受容器には、主に3つの種類がある。錐体は色に大きく反応する光受容器である。ヒトの場合、3種類の錐体が、短波長(青)、中波長(緑)、長波長(黄/赤)の一次反応に対応する[5]。杆体は、光の強弱に非常に敏感な光受容器で、薄暗い場所での視覚を可能にする。杆体と錐体の集中度や比率は、動物が昼行性か夜行性かに強く関連している。杆体と錐体の比率は、ヒトの場合で約20:1であるのに対し、モリフクロウのような夜行性動物では1000:1近くになる[5]。網膜神経節細胞は、交感神経反応に関与している。網膜に存在する約130万個の神経節細胞のうち、1-2%が感光性を持つと考えられている[6]。
視覚に関連する感覚ニューロンの問題や衰えは、次のような障害を引き起こす。
- 黄斑変性症 - 網膜と脈絡膜との間に細胞の残骸や血管が蓄積し、そこに存在するニューロンの複雑な相互作用が阻害あるいは破壊されることによる中心視野の変性[7]。
- 緑内障 - 網膜神経節細胞の障害で、視力低下から失明を引き起こす[8]。
- 糖尿病性網膜症 - 糖尿病による血糖コントロールの低下により、網膜の細い血管が損傷する[9]。
聴覚
編集聴覚系 (en:英語版) は、空気分子が振動することで発生する圧力波や音を、脳が解釈できる信号に変換する役割を担う。
この機械電気変換は、耳の中にある有毛細胞を介して行われる。その動きに応じて、有毛細胞は過分極または脱分極を起こすことができる。突出した不動毛にその動きが伝わると、Na+カチオンチャネルが開いてNa+が細胞内に流入し、その結果起こる脱分極によりCa++チャネルが開き、その神経伝達物質が求心性聴神経に放出される。有毛細胞には、内有毛細胞と外有毛細胞の2種類がある。内有毛細胞は感覚受容器である[10]。
聴覚系に関連する感覚ニューロンの問題は、次のような障害を引き起こす。
- 聴覚情報処理障害 - 脳内の聴覚情報が異常な状態で処理される。聴覚情報処理障害の患者は、通常、情報を得ることはできるものの、脳がそれを適切に処理できずに聴覚障害につながる[11]。
- 聴覚言語失認症 - 言葉の理解力は失われるが、聴力、会話、読み書きの能力は維持される。これは、側頭葉後上部[訳語疑問点]の損傷によって引き起こされ、やはり脳が聴覚入力を正しく処理することができなくなる[12]。
温覚
編集温度受容器は、温度の変化に反応する感覚受容器である。この受容器が機能する機構は不明であるが、最近の発見により、哺乳類には少なくとも2種類の温度受容器があることが明らかになった[13]。球状小体は皮膚受容体の一つで冷感感受性の受容体である。もう一つは、温感感受性受容体である。
機械受容器
編集機械受容器は、圧力や歪みなどの機械的な力に反応する感覚受容器である[14]。
機械受容器と呼ばれる特殊な感覚受容器細胞は、しばしば求心性線維を被包し、求心性線維をさまざまな種類の体性刺激に適合させるのを助ける。機械受容器はまた、求心性線維の活動電位発生の閾値を下げ、感覚刺激の存在下での発火を促進する[15]。
機械受容器の中には、膜が物理的に引き伸ばされたときに活動電位を発生させるものがある。
固有受容器(自己受容器とも)は、機械受容器のもう一つの種類であり、文字通り「自己のための受容器」を意味する。これらの受容器は手足やその他の身体部分に関する空間情報を提供する[16]。
侵害受容器は、痛みと温度変化を処理する責任を担っている。唐辛子を食べた後に経験する灼熱痛や刺激感(主成分であるカプサイシンによる)、メントールやイシリンなどの化学物質を摂取した後の冷感、そして一般的な痛みの感覚は、すべてこれらのニューロンと受容体の働きによるものである[17]。
機械受容器に問題は、次のような障害を引き起こす。
- 神経因性疼痛(神経障害性疼痛) - 感覚神経の損傷に起因する激しい痛みの状態[17]。
- 痛覚過敏 - 痛みに対する感受性の増加。知覚的なイオンチャネルであるTRPM8は、通常23-26度の温度に反応し、メントールやイシリンに関連する冷感を与える[17]。
- 幻肢症候群 - 存在しない手足に痛みや動きを感じる感覚系障害[18]。
内受容器
編集体内の変化に反応する内部受容器を内受容器(interoceptor)と呼ぶ[3]。
血液
編集大動脈小体と頚動脈小体には、末梢性化学受容器と呼ばれるグロムス細胞の集団があり、酸素濃度など血液中の化学的性質の変化を感知する[19]。これらは、多くの異なる刺激に反応するポリモーダルな受容器である。
侵害受容器
編集侵害受容器は、損傷を与える可能性のある刺激に反応し、脊髄と脳へ信号を送る。この過程は侵害受容(nociception)と呼ばれ、通常、痛みの感覚を認識させる[20][21]。侵害受容器は、「感知し、防御する」ために、体表だけでなく体内にも存在する[21]。侵害受容器は、損傷を与える可能性を示すさまざまな種類の侵害刺激を検出し、その刺激から逃れるための神経応答を開始する[21]。
中枢神経系とのつながり
編集頭部に存在する感覚ニューロンから来る情報は、脳神経を通って中枢神経系(CNS)に入る。対して、頭部よりも下に存在する感覚ニューロンからの情報は、脊髄に入り、31対の脊髄神経を通って脳へと向かう[23]。脊髄を通過する感覚情報は、明確に定義された経路をたどる。神経系は、どの細胞が活動しているかという観点から、感覚の違いをコード化している。
分類
編集適刺激
編集感覚受容器の適刺激(てきしげき)とは、その感覚受容器が適切な感覚伝達系を備えている刺激モダリティ(様式)のことである。適刺激は感覚受容器を分類するために用いられる。
- 圧受容器は、血管内の圧力に反応する。
- 化学受容器は、化学的刺激に反応する。
- 電磁波受容体は、電磁放射に反応する[24]。
- 電気受容体は、電場に反応する。
- ロレンチーニ器官は、電場、塩分、温度に反応するが、主に電気受容体として機能する。
- 水受容器は、湿度の変化に反応する。
- 磁気受容器は、磁場に反応する。
- 機械受容器は、機械的ストレスや機械的ひずみに反応する。
- 侵害受容器は、体組織の損傷や損傷の脅威に反応し、痛みを知覚するようになる(常にではなく、多くの場合)。
- 浸透圧受容器は、体液の浸透圧に反応する(視床下部など)。
- 固有受容器(自己受容器)は、位置感覚を提供する。
- 温度受容器は、熱さ、冷たさ、またはその両方の温度に反応する。
位置
編集感覚受容器は、その位置によって分類することができる。
形態
編集皮膚の表面近くにある体性感覚受容器は、通常、形態に基づいて2つのグループに分けられる。
- 自由神経終末は、侵害受容器と温度受容器を特徴づけるものであり、ニューロンの終末枝が無髄で、真皮と表皮全体に広がっているため、このように呼ばれる。
- 被包性受容体は、残りの種類の皮膚受容体で構成されている。カプセル化は、特殊な機能を果たすために存在する。
適応速度
編集- 緊張性受容器は、刺激に対してゆっくりと適応し[26]、刺激の持続時間にわたって活動電位を生成し続ける感覚受容器である[27]。このようにして、刺激の持続時間に関する情報を伝達する。いくつかの緊張性受容器は永続的に活性であり、バックグラウンドレベルを示す。そのような緊張性受容器の例には、疼痛受容器、関節包、および筋紡錘がある[28]。
- 相動性受容器は、刺激に対して急速に適応する感覚受容器である。細胞の反応は非常に速く減衰し、その後停止する[29]。刺激の持続時間に関する情報を提供しない代わりに[27]、それらのいくつかは、刺激の強度や速度の急激な変化に関する情報を伝える[28]。相動性受容器の例としてパチニ小体がある。
薬物
編集現在、感覚器系障害を処置または治療するために使用される数多くの薬物が市場に出回っている。たとえば、ガバペンチンは、非受容器ニューロン[訳語疑問点]に存在する電位依存性カルシウムチャネルの1つと相互作用することによって、神経因性疼痛を治療するために使用される薬物である[17]。薬によっては、他の健康問題と戦うために使用された場合でも、感覚器系に意図しない副作用を及ぼすことがある。聴器毒性薬は、アミノグリコシド系抗生物質のように、有毛細胞の働きを弱める毒素を使用して蝸牛に影響を与える薬物である。これらの毒素の使用により、K+ポンプ有毛細胞はその機能を停止する。その結果、聴覚信号伝達プロセスを駆動する蝸牛内電位によって生成されるエネルギーが失われ、難聴につながる[30]。
神経可塑性
編集エドワード・タウブによってシルバースプリング猿の脳における皮質再マッピングが観察されて以来、感覚系の神経可塑性に関する多くの研究が行われてきた。感覚系の障害の治療は大きく進歩した。タウブが開発した拘束運動療法のような技術は、感覚系に新しい神経回路を強制的に成長させることで、麻痺した手足を持つ患者が手足を再び使えるようにするのに役立った[31]。幻肢症候群とは、被切断者が切断された手足がまだ存在し、まだ痛みを感じていると認識する感覚器障害である。ヴィラヤヌル・S・ラマチャンドランが開発したミラーボックスは、幻肢症候群の患者が麻痺や痛みを伴う幻肢の知覚を軽減することを可能にした。これは、箱の中の鏡を使って、感覚系が片手ではなく両手を見ていると認める錯覚を作り出すことで、感覚系が「幻肢」を制御できるようにする単純な装置である。これにより、感覚系が切断された手足に徐々に順応し、この症候群を緩和することができる[32]。
ヒト以外の生物
編集流体力学的受容は、さまざまな動物種に備わっている機械受容の一形態である。(例: 魚類が水の動きを感知する側線と呼ばれる器官)
他の画像
編集-
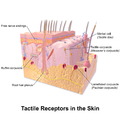 皮膚の触覚受容体
皮膚の触覚受容体 -
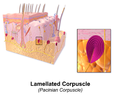 層板小体
層板小体 -
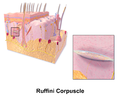 ルフィニ小体
ルフィニ小体 -
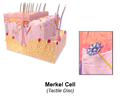 皮膚メルケル細胞
皮膚メルケル細胞 -
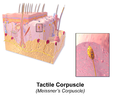 触覚小体
触覚小体 -
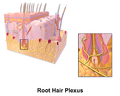 毛根神経叢
毛根神経叢 -
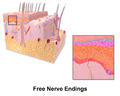 自由神経終末
自由神経終末







参照項目
編集脚注
編集- ^ Parsons, Richard (2018). CGP: A-Level Biology Complete Revision & Practice. Newcastle Upon Thynde: Coordination Group Publishing Ltd.. pp. 138. ISBN 9781789080261
- ^ Purves, Dale; Augustine, George; Fitzpatrick, David; Hall, William; LaMantia, Anthony-Samuel; McNamara, James; White, Leonard (2008). Neuroscience (4 ed.). Sinauer Associates, Inc.. pp. 207. ISBN 978-0878936977
- ^ a b Campbell, Neil (1996). Biology (4th ed.). Benjamin/Cummings Pub. Co. p. 1028. ISBN 0805319409
- ^ Breed, Michael D., and Moore, Janice. Encyclopedia of Animal Behavior . London: Elsevier, 2010. Print.
- ^ a b "eye, human." Encyclopædia Britannica. Encyclopædia Britannica Ultimate Reference Suite. Chicago: Encyclopædia Britannica, 2010.
- ^ Foster, R. G.; Provencio, I.; Hudson, D.; Fiske, S.; Grip, W.; Menaker, M. (1991). "Circadian photoreception in the retinally degenerate mouse (rd/rd)". Journal of Comparative Physiology A 169. doi:10.1007/BF00198171
- ^ de Jong, Paulus T.V.M. (2006-10-05). “Age-Related Macular Degeneration”. New England Journal of Medicine 355 (14): 1474–1485. doi:10.1056/NEJMra062326. ISSN 0028-4793. PMID 17021323.
- ^ Alguire, Patrick; Dallas, Wilbur; Willis, John; Kenneth, Henry (1990). “Chapter 118 Tonometry”. Clinical methods : the history, physical, and laboratory examinations (3 ed.). Butterworths. ISBN 978-0409900774. OCLC 15695765
- ^ “NIHSeniorHealth: Diabetic Retinopathy - Causes and Risk Factors”. nihseniorhealth.gov. 2017年1月14日時点のオリジナルよりアーカイブ。2016年12月19日閲覧。
- ^ Purves, Dale; Augustine, George; Fitzpatrick, David; Hall, William; LaMantia, Anthony-Samuel; McNamara, James; White, Leonard (2008). Neuroscience (4 ed.). Sinauer Associates, Inc.. pp. 327–330. ISBN 978-0878936977
- ^ “Auditory Processing Disorder (APD)”. British Society of Audiology APD Special Interest Group MRC Institute of Hearing Research. 2015年4月2日時点のオリジナルよりアーカイブ。2022年9月1日閲覧。
- ^ Stefanatos, Gerry A.; Gershkoff, Arthur; Madigan, Sean (2005-07-01). “On pure word deafness, temporal processing, and the left hemisphere”. Journal of the International Neuropsychological Society 11 (4): 456–470; discussion 455. doi:10.1017/S1355617705050538. ISSN 1355-6177. PMID 16209426.
- ^ Krantz, John. Experiencing Sensation and Perception Archived 2017-11-17 at the Wayback Machine.. Pearson Education, Limited, 2009. p. 12.3
- ^ Winter, R., Harrar, V., Gozdzik, M., & Harris, L. R. (2008). The relative timing of active and passive touch. [Proceedings Paper]. Brain Research, 1242, 54-58. doi:10.1016/j.brainres.2008.06.090
- ^ Purves, Dale; Augustine, George; Fitzpatrick, David; Hall, William; LaMantia, Anthony-Samuel; McNamara, James; White, Leonard (2008). Neuroscience (4 ed.). Sinauer Associates, Inc.. pp. 209. ISBN 978-0878936977
- ^ Purves, Dale; Augustine, George; Fitzpatrick, David; Hall, William; LaMantia, Anthony-Samuel; McNamara, James; White, Leonard (2008). Neuroscience (4 ed.). Sinauer Associates, Inc.. pp. 215–216. ISBN 978-0878936977
- ^ a b c d Lee, Y; Lee, C; Oh, U (2005). “Painful channels in sensory neurons”. Molecules and Cells 20 (3): 315–324. PMID 16404144.
- ^ Halligan, Peter W; Zeman, Adam; Berger, Abi (1999-09-04). “Phantoms in the brain”. BMJ: British Medical Journal 319 (7210): 587–588. doi:10.1136/bmj.319.7210.587. ISSN 0959-8138. PMC 1116476. PMID 10473458.
- ^ Satir, P. & Christensen, S.T. (2008) Structure and function of mammalian cilia. in Histochemistry and Cell Biology, Vol 129:6
- ^ Sherrington C. The Integrative Action of the Nervous System. Oxford: Oxford University Press; 1906.
- ^ a b c d e St. John Smith, Ewan (2017-10-14). “Advances in understanding nociception and neuropathic pain”. Journal of Neurology 265 (2): 231–238. doi:10.1007/s00415-017-8641-6. ISSN 0340-5354. PMC 5808094. PMID 29032407.
- ^ Zhao, Jianhua; Lin King, John V.; Paulsen, Candice E.; Cheng, Yifan; Julius, David (2020-07-08). “Irritant-evoked activation and calcium modulation of the TRPA1 receptor”. Nature 585 (7823): 141–145. Bibcode: 2020Natur.585..141Z. doi:10.1038/s41586-020-2480-9. ISSN 1476-4687. PMC 7483980. PMID 32641835.
- ^ Kalat, James W. (2013). Biological Psychology (11th ed.). Wadsworth Publishing. ISBN 978-1111831004
- ^ Michael J. Gregory. “Sensory Systems”. Clinton Community College. 2013年6月25日時点のオリジナルよりアーカイブ。2013年6月6日閲覧。
- ^ “Cutaneous receptor”. 2022年9月1日閲覧。
- ^ Binder, Marc D.; Hirokawa, Nobutaka; Windhorst, Uwe (2009). Encyclopedia of neuroscience ([Online-Ausg.]. ed.). Berlin: Springer. ISBN 978-3-540-29678-2
- ^ a b mentor.lscf.ucsb.edu/course/fall/eemb157/lecture/Lectures%2016,%2017%2018.ppt [リンク切れ]
- ^ a b “Sensory Receptor Function”. frank.mtsu.edu. August 3, 2008時点のオリジナルよりアーカイブ。2022年9月1日閲覧。
- ^ Sherwood, Lauralee; Klandorf, Hillar; Yancey, Paul (2012) (英語). Animal Physiology: From Genes to Organisms. Cengage Learning. ISBN 978-0840068651 13 December 2017閲覧。
- ^ Priuska, E.M.; Schacht, J. (1997). “Mechanism and prevention of aminoglycoside ototoxicity: Outer hair cells as targets and tools”. Ear, Nose, & Throat Journal 76 (3): 164–171. doi:10.1177/014556139707600310. PMID 9086645.
- ^ Schwartz and Begley 2002, p. 160; "Constraint-Induced Movement Therapy", excerpted from "A Rehab Revolution," Stroke Connection Magazine, September/October 2004. Print.
- ^ Blakeslee, Sandra; Ramachandran, V. S. (1998). Phantoms in the brain : probing the mysteries of the human mind. William Morrow & Company, Inc. ISBN 978-0688152475. OCLC 43344396
外部リンク
編集 ウィキメディア・コモンズには、感覚ニューロンに関するカテゴリがあります。
ウィキメディア・コモンズには、感覚ニューロンに関するカテゴリがあります。- 体性感覚受容器の主な種類